CUBIERTAS EXTRACELULARES
Sobre la membrana plasmática se pueden depositar secretados por las propias células, formando cubiertas extracelulares.
Las cubiertas extracelulares se corresponden con la matriz extracelular, presente entre las células de determinados tejidos animales, y con la pared celular eucariota, que rodea las células de hongos, algas y vegetales.
La matriz extracelular
Está formada por dos tipos de estructuras: láminas basales, que sirven de sostén bajo células epiteliales o alrededor de células musculares y adiposas; y matrices intercelulares en las que se encuentran las células de tejidos conectivos como el óseo, el cartilaginoso o el sanguíneo.
A. Composición de la matriz extracelular
Aunque su proporción varía en función del tipo celular, todas las matrices extracelulares están compuestas por proteínas y polisacáridos.
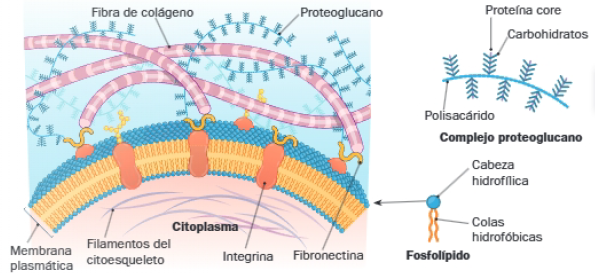
- Proteínas fibrosas: desempeñan funciones estructurales, que aportan consistencia y elasticidad a la matriz. La más frecuente, encargada de conferir resistencia a los tejidos animales es el colágeno. Entre las fibras elásticas destaca la elastina. Ambas proteínas suelen aparecer juntas y son especialmente abundantes en tejidos que sufren tracciones, como los de tendones y ligamentos.
- Glucoaminoglucanos (GAG), como el ácido hialurónico: aportan viscosidad a la matriz, pues atraen cationes que retienen agua. Forman una matriz gelatinosa que embebe las proteínas fibrosas, aportando soporte mecánico. La resistencia del cartílago se debe a su alto contenido de estos polisacáridos. Suelen unirse a una cadena polipeptídica central, originando proteoglucanos.
- Proteínas de adhesión, como la fibronectina y las lamininas: unen los componentes anteriores a las células, por lo que son responsables del ensamblaje de la matriz y favorecen la adhesión de las células entre sí. Se unen al colágeno y a los proteoglucanos, y a proteínas de la membrana plasmática llamadas integrinas. Gracias a estas uniones, las proteínas y los polisacáridos de la matriz quedan estrechamente asociados a la cara externa de la membrana, formando parte estos últimos glicocálix.
B.Funciones de la matriz extracelular
De las propiedades de los componentes anteriores derivan las funciones de las matrices extracelulares, entre las cuales destacan las siguientes:
- Participa en las interacciones célula-matriz, es decir, la propia matriz une las células de los tejidos a su lámina basal o matriz extracelular. Esto se debe a las uniones de sus proteínas de adhesión a las integrinas de membrana.; estas a su vez se unen al citoesqueleto, proporcionando estabilidad a las uniones entre célula y matriz.
- Dado que la matriz intercelular y la lámina basal mantienen unidas células de un mismo tejido o tejidos vecinos, contribuye al mantenimiento de los mismos e interviene en su organización durante el desarrollo embrionario y el crecimiento.
- Proporciona continuidad entre tejidos contiguos, estableciendo una vía de comunicación entre ellos, como ocurre entre el tejido del epitelio intestinal y el sanguíneo.
- Confieren resistencia y elasticidad en tejidos sometidos a esfuerzos mecánicos.
La pared celular vegetal
La pared celular vegetal se encuentra en torno a las células eucariotas vegetales. Está compuesta por polisacáridos, concretamente celulosa, hemicelulosa y pectinas, dispuestos en varias capas. Sus diversas funciones son fundamentales para el mantenimiento de la estructura de los vegetales, y de la adhesión y comunicación entre las células de sus tejidos.

A. Composición de la pared vegetal
Pese a que su cantidad varía en cada capa de la pared celular, estas están integradas por los siguientes componentes:
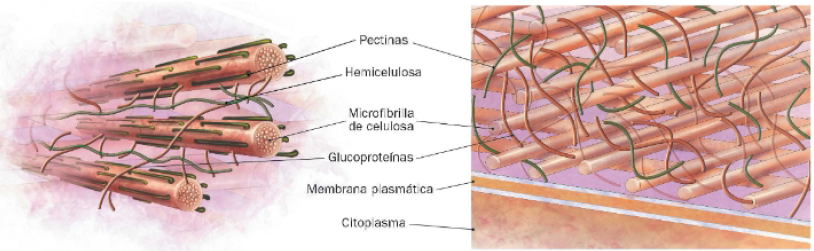
- Celulosa: se trata de un polímero lineal de glucosas, por lo que forma largas cadenas que se transportan al exterior celular tras ser sintetizadas en un complejo enzimático localizado en la membrana, la celulosa sintasa. La asociación de estas cadenas forma microfibrillas de celulosa que se disponen formando láminas. Es el componente más abundante.
- Hemicelulosa: este polisacárido es responsable de la resistencia mecánica de la pared celular, pues une estrechamente a las microfibrillas de celulosa para estabilizarlas.
- Pectina: gracias a la carga de su superficie, retiene moléculas de agua y forma una matriz de consistencia gelatinosa. En ella suele estar inmerso el conjunto formado por microfibrillas de celulosa y hemicelulosas, que le aportan soporte estructural. En una de las capas de la pared, la adición de glucoproteínas contribuye a esta función.
En las paredes celulares de determinados tejidos vegetales, los componentes anteriores pueden estar acompañados por otros polímeros (lignina, ceras, cutinas, suberina) e incluso sales.
B. Estructura de la pared celular vegetal
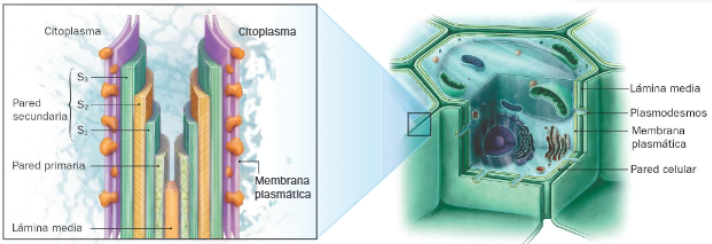
La pared celular vegetal es una estructura gruesa, formada por 2 o 3 capas en función del tipo de tejido o momento del desarrollo de la planta. Son las siguientes:
- Lámina media: se trata de la capa más externa, de modo que es compartida por células adyacentes de un tejido. Está formada principalmente por pectinas y es la primer que se deposita tras la división celular. Puede lignificarse tras la muerte celular.
- Pared primaria: compuesta por microfibrillas de celulosa entrelazadas al azar, cohesionadas por hemicelulosas y localizadas en la matriz de pectinas que, en esta capa, contiene glucoproteínas. Su flexibilidad es mayor en tejidos en crecimiento, para permitir su expansión.
- Pared secundaria: es la capa más próxima a la membrana plasmática, pero solo aparece en tejidos vasculares y de soporte tras haber cesado el crecimiento. En estos casos, aporta rigidez, pues contiene más celulosa que en el caso anterior y carece de pectinas. En ocasiones, se añaden otras substancias a su composición, como inclusiones de carbonato de calcio, lignina en las células del xilema y esclerénquima, ceras y cutina en las hojas, o suberina en raíces y tronco. Está formada por un número variable de estratos, en los que las microfibrillas se colocan paralelas pero cuya orientación varía entre capas.
C. Funciones de la pared celular vegetal
Las funciones de la pared celular vegetal son fundamentales para los vegetales:
- Une células de tejidos a través de la lámina media y posibilita su comunicación, gracias a la presencia de plasmodesmos, que son canales que interrumpen la continuidad de la pared, permitiendo la conexión y el intercambio de sustancias entre citoplasmas de células vecinas.
- Al ser rígida debido a la naturaleza y organización de sus componentes, define la forma celular y da soporte estructural a las plantas (permiten que se mantengan erguidas).
- Su rigidez evita que las células sufran lisis por la entrada osmótica de agua, que se produce debido a que el interior de la célula vegetal es hipertónico. Esta entrada de agua provoca que la célula esté turgente, lo que genera una presión sobre la pared, fundamental para el crecimiento de las células y para el soporte de las plantas (presión de turgencia).
- Actúa como barrera frente a agentes patógenos y, en los tejidos en los que presenta cutina y suberina, impermeabiliza la superficie de la planta evitando las pérdidas de agua.
MEMBRANA PLASMÁTICA
La estructura de las membranas celulares se basa en las propiedades de sus componentes principales, los fosfolípidos, que se disponen formando bicapas con proteínas asociadas.
Al observarla a través del microscopio electrónico de transmisión, dicha estructura se asemeja a las vías de un tren o a las capas de un sándwich, pues consiste en dos bandas oscuras separadas por una banda clara.

Todas las membranas eucariotas (la membrana plasmática y aquellas que limitan los orgánulos y el núcleo) presentan una organización común, y por ello recibe el nombre de membrana unitaria o unidad de membrana.
Composición de la membrana plasmática
La membrana plasmática está formada fundamentalmente por lípidos y proteínas, y aunque su proporción varía entre distintos tipos de células, cada uno de estos componentes representa en torno al 50% del peso de la misma. Los glúcidos son un componente minoritario, y aparecen unidos a los anteriores formando glucolípidos y glucoproteínas.
A. Lípidos de membrana
Los lípidos desempeñan funciones estructurales en la membrana y son de 3 tipos:
- Fosfolípidos: son los lípidos más abundantes, pues originan las bicapas que constituyen la estructura básica de las membranas. Esto se debe a su carácter anfipático, mientras que las cabezas polares interaccionan con los medios acuosos intracelular y extracelular, sus colas apolares encuentran resguardo en el interior de la bicapa fosfolipídica resultante.
- Esteroles: como el colesterol, que se encarga de estabilizar la membrana, pues, al insertarse en la bicapa de fosfolípidos, aumenta su rigidez.
- Glucolípidos: aparecen en la cara externa de la membrana (hacia el medio extracelular) y son el resultado de la unión covalente entre un fosfolípido y un oligosacárido.
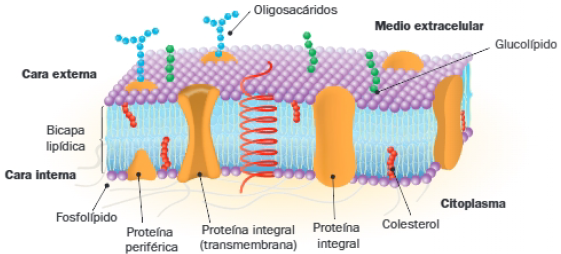
B. Proteínas de membrana
Las proteínas de membrana son las responsables de las funciones específicas de la membrana plasmática, a la cual se unen a ella siendo hasta 100 veces más voluminosas que los fosfolípidos que la conforman.
Según presenten mayor o menor grado de asociación con la membrana plasmática, las proteínas de membrana serán integrales o periféricas
- Proteínas integrales: presentan un dominio que se inserta en la bicapa lipídica, y otro que sobresale por la cara externa de la membrana, por la interna o por ambas (proteínas transmembranales). Su unión con la membrana es fuerte, mediante enlaces hidrófobos con su interior apolar, por lo que también se denominan proteínas intrínsecas.
- Proteínas periféricas: a diferencia de las anteriores, no está embebidas en el interior hidrofóbico de las bicapas. Se unen mediante enlaces iónicos a proteínas integrales, pero también pueden unirse a fosfolípidos. Estas uniones son más débiles y por ello reciben el nombre de proteínas extrínsecas.
La mayoría de proteínas que asoman a la cara externa de la membrana están unidas covalentemente a oligosacáridos, dando lugar a glucoproteínas.
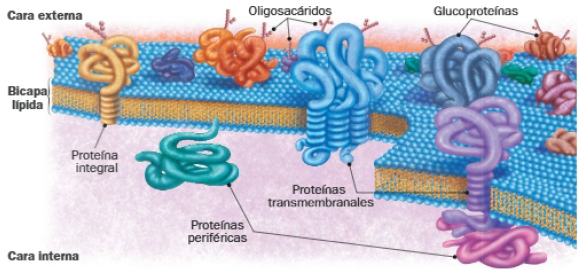
C. Glúcidos de membrana
Los glúcidos de membrana se corresponden con los restos glucídicos que se unen a fosfolípidos y proteínas de la cara externa de la membrana, esto es, con los glucolípidos y las glucoproteínas. Estas fracciones glucídicas, junto con las que forman parte de las matrices extracelulares (GAG), conforman el glicocálix, una capa glucídica que reviste la superficie de la célula.
El glicocálix o glucocáliz interviene en diversas funciones:
- Interacción entre células, o entre células y matrices extracelulares.
- Reconocimiento entre células, ya que aporta propiedades antigénicas.
- Reconocimiento de sustancias durante los procesos de fagocitosis y pinocitosis.
- Protección de la superficie celular contra posibles lesiones.
- Formación de una barrera contra microorganismos patógenos.
- Aporta viscosidad, importante para el movimiento de ciertas células.
La presencia de oligosacáridos en la cara externa de la membrana hace que se puedan diferenciar ambas caras por su composición. Además, el colesterol es más abundante en la cara externa, y las proteínas periféricas en la cara interna de las membranas. Es por ello que decimos que las membranas son asimétricas en cuanto a su composición.
Estructura de la membrana plasmática
Los primeros modelos de la estructura de la membrana la consideraron un sólido rígido, cuya bicapa estaría cubierta superior e inferiormente por una capa de proteínas.
A. Modelo del mosaico fluido
Según este modelo, la membrana se corresponde con un fluido bidimensional, pues está formada por moléculas con libertad de movimiento dispuestas en dos capas.
- Debido a su carácter anfipático, los fosfolípidos de membrana se disponen formando dos capas (bicapa lipídica), a las que, de forma asimétrica, se unen moléculas de colesterol, oligosacáridos y proteínas. Las regiones polares de dichas moléculas se sitúan en ambas caras de la membrana, y las apolares quedan resguardadas en su interior.
- Dichas uniones son flexibles, pues ciertas moléculas (fosfolípidos y proteínas) pueden moverse individualmente en el interior de la bicapa, haciendo que se comporte como un fluido. El movimiento más frecuente es la difusión lateral de dichas moléculas, aunque los fosfolípidos también pueden experimentar rotaciones, flexiones o movimientos flip-flop.
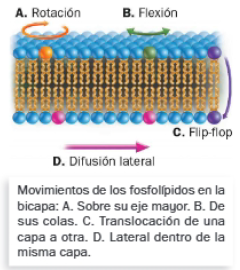
Dicho esto, se entiende que la fluidez de la membrana es fundamental, pues de ella dependen muchas funciones.
La fluidez de la membrana aumenta cuando la temperatura se aproxima al punto de fusión de los lípidos y con el grado de insaturación de los ácidos grasos de los fosfolípidos. La longitud y la saturación de sus cadenas, así como la proporción de colesterol, aumentan la rigidez de la membrana.
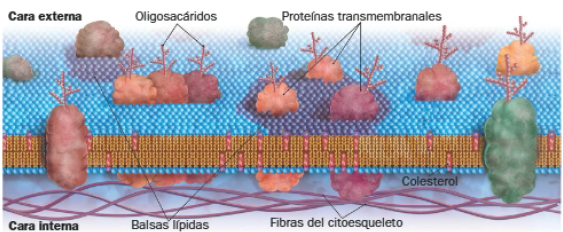
Sin embargo, determinadas moléculas de las membranas, como proteínas con funciones específicas, deben mantener su posición para garantizar el correcto funcionamiento de la célula. Esto es posible gracias a la existencia de balsas lipídicas, placas semisólidas formadas por asociaciones de colesterol con proteínas y fosfolípidos que limitan sus movimientos. Esto genera dominios funcionales en la membrana, fundamentales en células polarizadas (aquellas en las que la superficie apical desempeña una función diferente a la superficie a la superficie basal). La interacción de dichas proteínas con el citoesqueleto contribuye a estabilizar las balsas lipídicas.
La siguiente imagen resume el modelo actual del mosaico fluido.
Transporte a través de la membrana plasmática
La membrana plasmática envuelve la célula sin generar un límite hermético, pues a través de ella se realizan intercambios de sustancias entre el citoplasma y el medio extracelular. Las sustancias para transportar pueden ser pequeñas o formas complejos macromoleculares, lo que determina el mecanismo de transporte mediante el que cruzan la membrana.
A. Mecanismos de transporte iónico y molecular
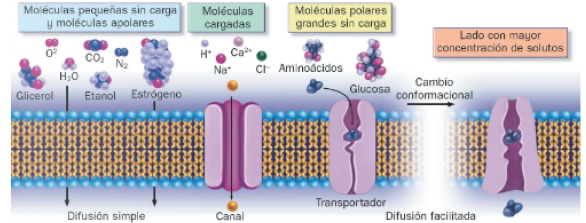
La membrana es permeable a pequeñas moléculas sin carga y a moléculas hidrofóbicas o apolares, que atraviesan directamente la bicapa de lípidos. Sin embargo, es impermeable a moléculas cargadas, como iones y a la mayoría de moléculas polares, como las biomoléculas orgánicas. Debido a su carga o que son hidrosolubles, no pueden interaccionar con las colas hidrofóbicas de los fosfolípidos, y por ello son transportadas por proteínas de membrana denominadas proteínas de transporte.
Dado que ciertas moléculas pueden difundir a través de la bicapa lipídica y otras no, la membrana plasmática es semipermeable o parcialmente permeable.
El dominio transmembrana de las proteínas de transporte permite el paso de moléculas incapaces de cruzar directamente la bicapa. Existen dos tipos de proteínas de transporte: los canales proteicos o proteínas de canal, y las proteínas transportadoras o transportadores de membrana. A pesar de que existen canales no selectivos, la existencia de proteínas de transporte determina la permeabilidad selectiva de las membranas, pues transportan moléculas específicas, mantenimiento la composición interna de la célula.
En función de si intervienen o no proteínas de transporte, y si el funcionamiento de estas consume energía, se diferencian varias modalidades de transporte de iones y moléculas.
Transporte pasivo o difusión
El transporte pasivo no requiere gasto energético, pues ocurre de forma espontánea a favor de gradiente de concentración. Por lo tanto, las moléculas se mueven en el sentido que determina su gradiente, atravesando la membrana de uno de los siguientes modos:
- Difusión simple: consiste en cruzar la bicapa lipídica directamente, interaccionando con los fosfolípidos. Muy pocas moléculas pueden difundir libremente a través de las membranas, pues deben ser pequeñas, sin carga e hidrofóbicas. Ciertas moléculas polares pueden hacerlo, además de neutras, deben presentar un tamaño reducido.
- Difusión facilitada: las moléculas difunden a través de canales proteicos o proteínas transportadoras. Entre los primeros se encuentran los canales iónicos, conductos que permiten el paso de iones (sodio, potasio y cloro). Las segundas transportan moléculas, como la glucosa, al sufrir cambios conformacionales ocasionados por la unión de las mismas.
Transporte activo
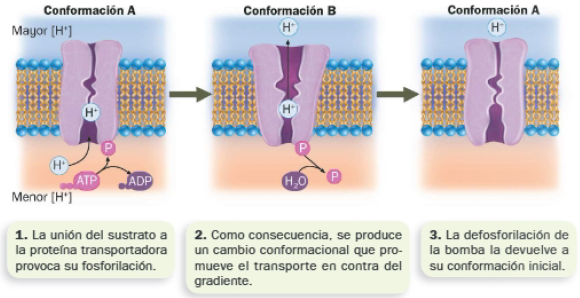
El transporte activo consume energía, pues ocurre en contra del gradiente de concentración o carga. Cuando se trata de iones o moléculas que no pueden difundir a través de la membrana, el transporte lo llevan a cabo proteínas transportadoras denominadas bombas, que consumen la energía liberada en una reacción de hidrólisis de ATP acoplada al transporte (tienen actividad ATPasa).
Entre los transportadores se encuentran las bombas iónicas, encargadas de mantener diferencias de concentración de ciertos iones dentro y fuera de la célula (gradiente iónico). El gradiente iónico a través de la membrana plasmática es fundamental, no solo para mantener el potencial de membrana necesario para la propagación de los impulsos nerviosos, sino para mantener el volumen celular (el transporte de iones en contra de gradiente evita la entrada osmótica de agua al medio intracelular, hipertónico).
Sentido del transporte
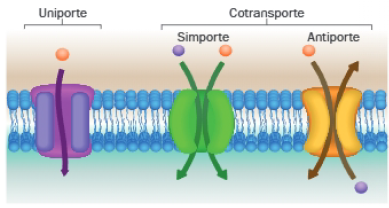
Los tipos de transporte también se pueden clasificar en función del número de moléculas transportadas y del sentido del transporte. En función de ello, hablaremos de uniporte o cotransporte, el cual puede ser de tipo simporte o antiporte.
B. Mecanismos de transporte macromolecular
El transporte de moléculas de alto peso molecular lleva consigo un gasto de energía y el tránsito de vesículas a través de la membrana celular, pequeñas bolsas membranosas que portan las sustancias que se deben movilizar.
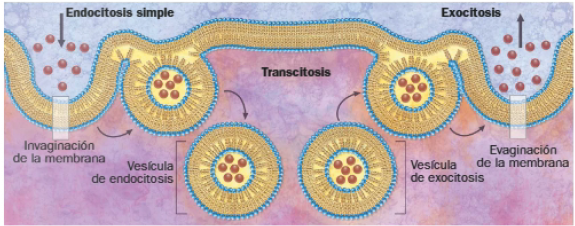
Los mecanismos de endocitosis introducen macromoléculas en la célula, al provocar la formación de vesículas por invaginación de la membrana plasmática. La exocitosis sigue el sentido opuesto, y expulsa sustancias gracias a la fusión de la membrana de las vesículas de exocitosis con la membrana celular. Un equilibrio entre ambos procesos asegura el mantenimiento del volumen de la misma.
Endocitosis
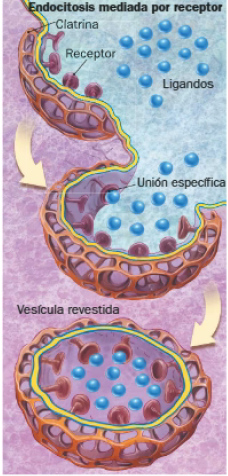
Existen dos tipos de endocitosis: simple y mediada por receptor. Por medio de esta última se transportan moléculas especialmente relevantes para la célula, como hierro, insulina o colesterol. Se unen especialmente a receptores presentes en la membrana celular y son englobadas en una vesícula revestida por una proteína denominada clatrina.
Una vez en el interior de la célula, las vesículas de endocitosis se unen a ciertos componentes celulares con uno de los siguientes fines:
- Digestivos: las vesículas se fusionan con lisosomas, originando vacuolas digestivas. Cuando las partículas ingeridas se encuentran en una fase líquida, se denomina pinocitosis. La ingestión de partículas sólidas mayores recibe el nombre de fagocitosis, que lleva consigo la formación de pseudópodos y de vesículas más voluminosas, los fagosomas.
- De transporte intracelular: el contenido de las vesículas debe llegar a un orgánulo concreto de la célula o a otra célula vecina. Para ello, se fusiona con la membrana de un orgánulo o de su propia célula, donde iniciará el proceso de exocitosis. Este tipo de tránsito intracelular se denomina transcitosis y ocurre, por ejemplo, en las células endoteliales.
Exocitosis
La fusión de la membrana externa e interna de las vesículas de exocitosis con la cara interna y externa de la membrana, respectivamente, genera un orificio a través del que se libera el contenido vesicular, con alguna de las siguientes funciones:
- Estructural (secreción constitutiva): liberación de sustancias sintetizadas en la célula que deben añadirse a sus cubiertas. (matriz extracelular, pared, membrana o glicocálix).
- De relación (secreción regulada): la célula segrega moléculas en respuesta a estímulos externos, tales como hormonas, neurotransmisores o enzimas digestivas.
- De excreción: eliminación de productos de deshecho generados en procesos de digestión celular, como la fagocitosis.
Receptores de membrana y señalización celular
La señalización celular o comunicación célula-célula abarca un conjunto de procesos que permiten el intercambio de señales entre las células de un ser vivo, con el fin de coordinar su actividad individual para que el organismo funcione correctamente como un todo.
Esto es posible gracias a la existencia de moléculas señalizadoras, secretadas o expresadas en la superficie celular, y de unas proteínas de membrana o citoplasmáticas denominadas receptores. Las comunicaciones intercelulares se desarrollan en las siguientes etapas:
- Recepción de un estímulo interno o externo por una célula receptora, que lo transforma en una célula señal (hormonas, fitohormonas, factores de transcripción, neurotransmisores).
- Unión específica de la molécula señal al receptor adecuado, localizado en otra célula (célula diana), en la que se desencadenan una serie de eventos moleculares. Cuando el receptor se localiza en la superficie de la célula, suele generarse una reacción en cascada, en la que intervienen los segundos mensajeros, como el AMPc.
- Respuesta al estímulo inicial, que puede inhibir o estimular diversos procesos celulares (metabólicos, de crecimiento, de diferenciación, de movimiento, de expresión génica).
Este tipo de comunicación es fundamental en organismos pluricelulares, en los que existe una gran variedad de vías de señalización que controlan su supervivencia y desarrollo.
Diferenciaciones de la membrana plasmática
Las diferenciaciones de la membrana plasmática consisten en modificaciones de la membrana plasmática que le permiten desempeñar funciones concretas. De este modo, puede replegarse hacia el interior para generar regiones en las que ocurren funciones concretas (invaginaciones, como las de ciertas células renales), o prolongarse para formar estructuras digitiformes que aumentan la superficie celular (microvellosidades). Los estereocillos son otras prolongaciones de membrana, cuyo movimiento es transformado en un impulso nervioso, como ocurre en las células encargadas de la audición y del equilibrio.
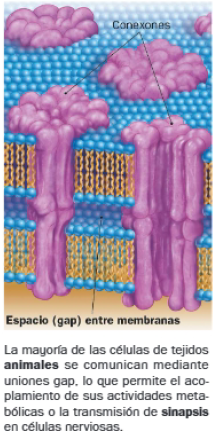
A. Uniones tipo gap
Las uniones de tipo gap son canales formados por un conjunto de proteínas transmembranales, cuya disposición forma un poro en el interior de una estructura cilíndrica (conexón). Los conexones de células contiguas están alineados, permitiendo el intercambio de iones y pequeñas moléculas entre sus citoplasmas. El dominio extracelular de estas proteínas genera un espacio (gap) entre las membranas de las células adyacentes.
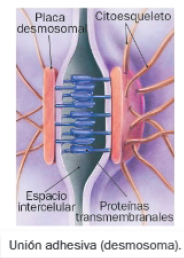
B. Uniones adhesivas
Las uniones adhesivas son uniones mecánicas entre células. Entre las de tipo estable destacan los desmosomas, que establecen uniones célula-célula, y los hemidesomas, responsables de uniones célula-matriz. En el caso de los desmosomas, las proteínas transmembranales de células vecinas se conectan entre sí, y al citoesqueleto de la célula de la que proceden mediante las proteínas de la placa desmosomal. Los hemidesmosomas reciben esta denominación porque las proteínas transmembranales se conectan con elementos de la matriz extracelular contigua, y no con las proteínas de membrana de otra célula.
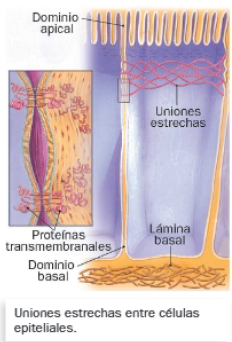
C. Uniones estrechas
Son uniones íntimas entre células, pues no dejan espacio entre ellas. Su finalidad es impedir el intercambio de sustancias entre células adyacentes o entre dominios de células polarizadas, como las del epitelio intestinal.
Las proteínas transmembrana de las diferentes células se unen a lo largo de una banda que rodea toda la célula, originando una red proteica que actúa como una barrera al paso de sustancias. Es habitual que, próximas a estas uniones, aparezcan otras de tipo adhesivo, para reforzar la unión intercelular.
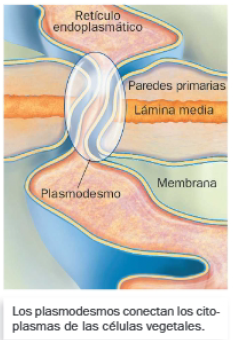
D. Plasmodesmos
Son canales que aparecen en la pared celular vegetal y su funcionamiento es análogo al de las uniones gap, pues permiten intercambios entre citoplasmas de células vecinas. Dicha comunicación se establece por medio de interrupciones en la pared formadas durante su síntesis y suelen estar atravesadas por membranas de retículo endoplasmático liso.
Tienen un papel clave en procesos de regulación, pues permiten el paso de pequeñas moléculas, como factores de transcripción o ARN.
Funciones de la membrana plasmática
- Su función principal es delimitar la célula, sin que esto signifique originar un límite pasivo, sino una barrera selectiva al paso de diversas sustancias. Las características de sus fosfolípidos y proteínas de transporte le permiten regular los intercambios de moléculas entre la célula y su entorno, lo que determina la composición del citoplasma.
- Muchas de sus moléculas proporcionan identidad celular, por lo que interviene en procesos de reconocimiento celular.
- Control de gradientes de concentración y electroquímicos gracias a las proteínas transportadoras, a los que se puede acoplar la fosforilación oxidativa (síntesis de ATP).
- Participa en procesos de nutrición, transporte intracelular, secreción y excreción llevados a cabo por endocitosis y exocitosis.
- Participa en procesos relación, con el exterior o entre células, gracias a la presencia de receptores, que captan señales y posibilitan la elaboración de respuestas.
- Dado que puede sufrir diferenciaciones, puede realizar funciones específicas y meditar procesos de adhesión y comunicación celular, fundamental en organismos pluricelulares.
- Aporta puntos de anclaje al citoesqueleto, que contribuyen a mantener la estructura celular.
NÚCLEO CELULAR
En el transcurso de la evolución, el núcleo de células eucariotas se forma como consecuencia de la compartimentación del material genético, al verse rodeado por membranas intracelulares que lo separan del resto del contenido citoplasmático. Debido a ello, desempeña funciones relacionadas con el control de la célula.
Su presencia es una característica definitoria de las células eucariotas y su estructura varía en función del momento del ciclo de vida de las mismas.
El núcleo interfásico alude al aspecto del núcleo durante la interfase, etapa del ciclo celular en la que la célula crece y realiza su función normal. Cuando la interfase está llegando a su término y la célula se prepara para su división, el núcleo comienza a sufrir cambios estructurales y pasa a denominarse núcleo en división.
Estructura del núcleo interfásico
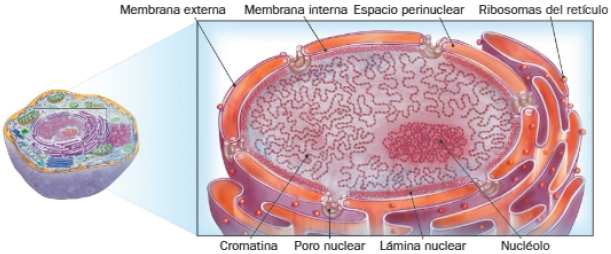
El núcleo celular está delimitado por una envoltura nuclear, formada por una membrana doble perforada por poros nucleares. Su interior está ocupado por un nucleoplasma, matriz en la que está inmerso el ADN y proteínas implicadas en su metabolismo.
En la interfase, los cromosomas se encuentran en forma de cromatina o estado relajado del ADN. Es decir, en lugar de la forma compacta o altamente condensada que adquieren antes de la división celular, en esta fase están descondensados. Sin embargo, para que las largas hebras de ADN quepan en el reducido diámetro nuclear, están ligeramente plegadas gracias a la asociación con unas proteínas denominadas histonas.
Al observar el núcleo interfásico al microscopio electrónico de transmisión, se diferencia una región de cromatina más oscura o electrodensa, el nucléolo.
A. Envoltura nuclear y poros nucleares
La envuelta nuclear separa el ADN del resto de la célula, generando un compartimento metabólico independiente en el que transcurren los procesos relacionados con el ADN y su regulación. Es por ello que muchas de las proteínas del nucleoplasma se corresponden con enzimas involucradas en estos procesos.
Otra de las funciones la envuelta nuclear es actuar como barrera selectiva al paso de sustancias entre núcleo y citoplasma, que son transportadas a través de las membranas de la propia envoltura y de los poros nucleares.
Además, se encarga de mantener la posición del núcleo en la célula, pues establece uniones con el citoesqueleto.
Componentes de la envoltura nuclear
Está formada por dos membranas concéntricas, las membrana nuclear externa y la membrana nuclear interna, separadas por un espacio intermembranoso denominado espacio perinuclear.
Dichas membranas se corresponden con bicapas lipídicas, que contienen proteínas unidas externamente al citoesqueleto e internamente a la llamada lámina nuclear. Esta está formada por una red de proteínas fibrosas que interacciona con la cromatina, aportándole puntos de anclaje, y con la envoltura nuclear, pues regula su desintegración y crecimiento antes y después de la división celular.
Por su parte, la membrana celular externa se continúa con las membranas del retículo endoplasmático, motivo por que se pueden encontrar ribosomas adheridos a la superficie citoplasmática del núcleo. En consecuencia, el espacio perinuclear está conectado con el lumen del retículo.
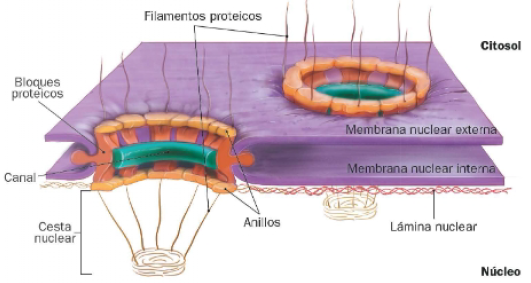
Estructura de los poros nucleares
Las membranas nucleares interna y externa se fusionan en algunos puntos, originando los poros nucleares. En estos orificios si sitúan los denominados complejos del poro nuclear, complejos proteicos de gran peso molecular en cuyo interior se localiza un canal que regula el paso de moléculas incapaces de difundir a través de las membranas de la envuelta nuclear.
Cada complejo está formado por ocho bloques proteicos, dispuestos en el interior de los poros nucleares formando una estructura octogonal. Este se une a dos anillos, citoplasmáticos y nuclear, desde los que se extienden filamentos de proteínas. Estos filamentos forman una estructura característica, la cesta nuclear, hacia el interior del núcleo. Dichas proteínas, y las situadas en el canal, interaccionan con las sustancias que se van a transportar.
B. Nucléolo
Aunque su tamaño varía en función de la actividad transcripcional, el nucléolo es un cuerpo denso visible en el interior del núcleo interfásico, como si de un pequeño núcleo se tratase. Su número también puede variar, pues las neuronas suelen presentar más de uno.
En esta zona del núcleo ocurren las síntesis de ARN ribosómicos y su ensamblaje con las dos subunidades que forman los ribosomas eucariotas. Por ello, el ADN que forma nucléolo se corresponde con la cromatina que codifica el ARNr. Estos genes forman parte de diferentes cromosomas, y su conjunto constituye el organizador nucleolar (NOR).
En la estructura del nucléolo se diferencian zonas menos densas y zonas con aspecto granular, formadas por ARN y proteínas. Las primeras se corresponden con los lugares en los que ocurre la transcripción del ADN nucleolar, o síntesis de ARNr; las segundas, con los lugares donde estos se ensamblan con las subunidades ribosómicas. Dichas subunidades están formadas por proteína, por lo que se sintetizan en el citoplasma (ribosomas). Después de ello, entran en el núcleo de sus poros, se unen al ARNr en las zonas granulares del nucléolo y regresan al citoplasma para ensamblarse y originar un ribosoma 80S.
Funciones del núcleo celular
Al contener la información genética, se corresponde con el centro de control celular, ya que se encarga de la transmisión de la herencia genética y de la expresión de los genes. Se debe a que en su interior ocurren los procesos de replicación y transcripción del ADN, así como la maduración del ARN sintetizado en este proceso. También interviene en la división celular.
El hecho de que la envuelta nuclear permita el paso de ciertas moléculas en lugar de limitar un compartimento estanco, resulta clave para el transcurso de los procesos anteriores, que necesitan y generan moléculas que viajan entre el núcleo y el citoplasma (ARN, proteína, enzimas, nucleótidos o subunidades ribosómicas).
Otras características del núcleo
A pesar de que los eritrocitos de mamíferos son células enucleadas (pierden su núcleo al diferenciarse para realizar su función), la mayoría de las células de los organismos pluricelulares presentan un núcleo. Con todo, células como los hepatocitos o los paramecios son binucleadas, y las células que forman plasmodios o sincitios son plurinucleadas.
Aunque la posición que ocupa en la célula suele ser central, en algunas ocasiones el núcleo aparece desplazado por otras estructuras, como la gran vacuola en las células vegetales o los acúmulos de grasa en los adipocitos.
La forma del núcleo puede variar pues, pese a que predominan las formas esféricas, también pueden ser fusiformes o polilobulados, como en el caso de los neutrófilos.
En cuanto al tamaño nuclear, permanece constante para cada tipo celular, y suele oscilar muy poco. Este valor de oscilación depende de la función del tipo de célula en cuestión, siendo mayor, por ejemplo, en la célula embrionaria que en un espermatozoide. Además, el tamaño del núcleo es directamente proporcional al tamaño del citoplasma, y ocupa sobre un 10% del volumen celular.
Fuente: Biología 2 (Mc Graw Hill)