COMPONENTES DEL CITOPLASMA
Las células eucariotas presentan una serie de elementos comunes: membrana plasmática, núcleo y citoplasma. El componente acuoso del citoplasma se denomina citosol, y en él se encuentran inmersos el citoesqueleto y los orgánulos celulares. Además, puede presentar inclusiones citoplasmáticas de diversa naturaleza.
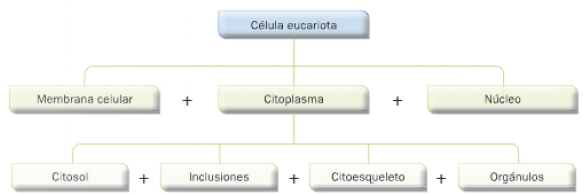
Citosol (Hialoplasma)
Es una matriz líquida formada fundamentalmente por agua, en la que se encuentran diversos solutos como enzimas, soluciones tampón, iones y moléculas orgánicas (glúcidos, aminoácidos, ARN y ATP).
Aquí vamos a sintetizar las principales funciones del citoplasma:
- Interviene en la regulación del pH intracelular, pues contiene tampón fosfato.
- Es el medio donde ocurren numerosas reacciones metabólicas, llevadas a cabo por las diversas enzimas que contiene.
- Puede variar su viscosidad.
- En ciertas células, almacena sustancias en forma de acúmulos o inclusiones.
Inclusiones citoplasmáticas
Se encuentran en el citosol y se corresponden con acumulaciones de sustancias inertes e hidrófobas.
Algunos de estos tipos de inclusiones son:
- Inclusiones cristalinas: suelen ser de naturaleza proteica, y pueden encontrarse, además de en el citoplasma, en alguno de los compartimentos celulares. Así, pueden formarse inclusiones cristalinas en el interior del núcleo o de la vacuola vegetal.
- Inclusiones de reserva: de glúcidos o lípidos. Entre las primeras destacan las inclusiones de polisacáridos, almidón de ciertas células vegetales, y gránulos de glucógeno de hepatocitos y fibras musculares. Entre las segundas, las inclusiones de triglicéridos de los adipocitos.
- Pigmentos: predominan las inclusiones de melanina en los melanocitos, responsables de la coloración oscura de la piel; y de carotenoides en las células de frutos de color anaranjado.
CITOESQUELETO
Está consituido por una red proteica tridimensional que se extiende por el citoplasma. Los componentes del citoesqueleto son los microtúbulos, los microfilamentos de actina y los filamentos intermedios, cuyo grosor se sitúa entre los dos anteriores. Los dos primeros son los más abundantes y se caracterizan por su comportamiento dinámico, mientras que los filamentos intermedios realizan funciones estáticas, estructurales.
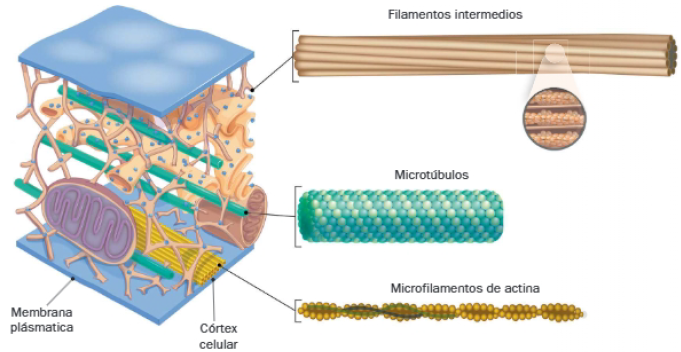
Aquí vamos a enumerar las principales funciones del citoesqueleto:
- Soporte y estabilidad mecánica de la célula y del propio citoesqueleto.
- Movimiento intracelular.
- Posicionamiento de los elementos celulares (orgánulos y núcleo).
- Determinación de la forma celular.
- Forma el esqueleto interno de centriolos, cilios, flagelos, microvellosidades y estereocilios.
- Formación del huso mitótico durante la división celular.
- Interviene en otras funciones como la contracción muscular, el crecimiento de la pared celular y en las interacciones entre células, y entre éstas y la matriz extracelular.
Componentes del citoesqueleto
A. Microtúbulos
Los microtúbulos son los elementos del citoesqueleto más gruesos. Su estructura tubular deriva de la polimerización de una proteína globular denominada tubulina.
Los microtúbulos están constituidos por 13 protofilamentos, elementos lineales formados por la sucesión de dímeros de tubulina, cuya disposición cilíndrica deja un núcleo central hueco. Al tratarse de estructuras dinámicas, su longitud varía con gran facilidad, por lo que las reacciones de polimerización y la hidrólisis de tubulina, que dependen del GTP, se alternan continuamente para satisfacer las necesidades de la célula.
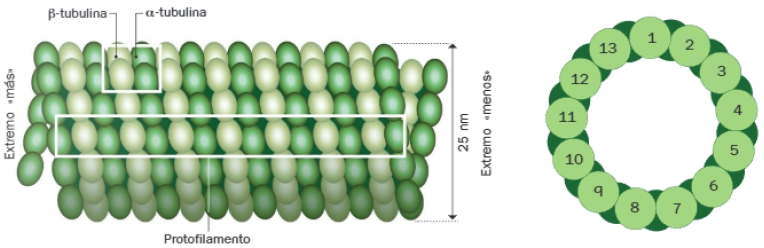
Debido a que los microtúbulos pueden sufrir rápidas remodelaciones, desempeñan funciones dinámicas, como la participación en movimientos celulares. En muchos de ellos intervienen proteínas motoras, cuyo desplazamiento a lo largo de los microtúbulos contituye un motor molecular que transporta elementos hacia el centro o la periferia de la célula, respectivamente. Existen otras proteínas que se asocian a los microtúbulos, como las MAPS, que regulan su comportamiento dinámico.
Aquí vamos a destacar las principales funciones de los microtúbulos:
- Transporte intracelular mediado por dineínas y quinesinas, de macromoléculas y elementos celulares, como vesículas de transporte o ciertos orgánulos (mitocondrias y cloroplastos).
- Organización intracelular: con la ayuda de las proteínas motoras, se encargan de posicionar los orgánulos membranosos en el citoplasma.
- Determinan la dirección de crecimiento de las paredes celulares, pues orientan el movimiento de la celulosa sintasa, y provocan que las microfibrillas de celulosa se dispongan paralelas a los microtúbulos corticales.
- Intervienen en el mantenimiento de la forma de determinadas células, como los eritrocitos.
- Movimiento y separación de cromosomas durante la división celular, pues son los elementos estructurales de huso mitótico.
- Cuando están presentes en el centrosoma, forman la estructura de los centriolos.
- Intervienen en determinados tipos de locomoción celular, ya que forman el esqueleto interno de cilios y flagelos.
Centros organizadores de microtúbulos
En las células animales, los microtúbulos se ensamblan a partir de los denominados centros organizadores de microtúbulos (COMT), que en ellas se corresponden con el centrosoma.
Por su parte, las células vegetales carecen de centriolos y de centros organizadores de microtúbulos. Sus microtúbulos se encuentran bajo la membrana plasmática y forman una matriz desde la que desempeñan su función principal: guiar la síntesis de la pared celular.
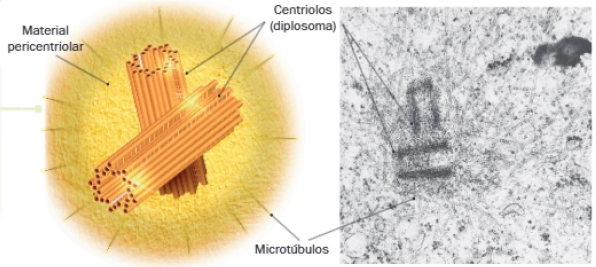
Cada celula animal contiene un centrosoma formado por:
- Dos centriolos dispuestos perpendicularmente (diplosoma).
- Material pericentriolar: material amorfo que rodea los centriolos.
El material pericentriolar es el responsable de iniciar la polimerización de los microtúbulos, hecho que determina su polaridad: mientras que el extremo que permanece anclado al COMT recibe la denominación de extremo negativo, el que sufre adición o disociación de dímeros de tubulina se corresponde con el extremo positivo.
Los centriolos son estructuras no membranosas presentes en el centrosoma de células animales y en la base de cilios y flagelos, e intervienen en la organización de microtúbulos.
Los centriolos están formados por 9 tripletes de microtúbulos que se disponen en torno a un núcleo central formando un cilindro. Esta zona central está ocupada por fibrillas radiales que se unen en un eje central, a modo de rueda de carro.
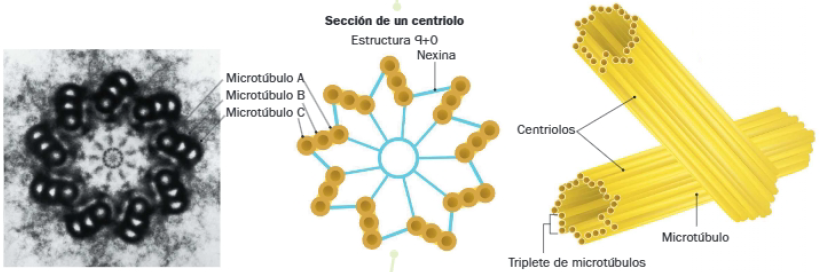
En cada triplete, el microtúbulo A se corresponde con el más interno y el C con el más externo. El B presenta una posición intermedia. Mientras que el microtúbulo A es completo, el B y el C son incompletos, pues comparten 3 de sus 10 protofilamentos con los microtúbulos anteriores. Los tripletes están conectados por medio de una proteína denominada nexina, que une los microtúbulos A y C de tripletes contiguos.
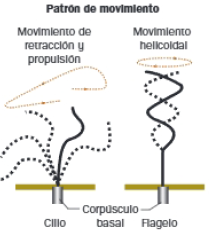
Cilios y flagelos
Son apéndices celulares cuya estructura interna, formada por microtúbulos, se encuentra por la membrana plasmática y consta de los siguientes elementos:
- Corpúsculo basal: deriva de un centriolo que se localiza bajo la membrana plasmática, por lo que sus microtúbulos se organizan siguiendo la estructura 9+0. Constituye el anclaje de los apéndices e inicia el crecimiento de los microtúbulos axonemales.
- Raíces ciliares: conjunto de microfilamentos situados bajo el corpúsculo basal, que se encarga de coordinar el movimiento de los cilios.
- Tallo o axonema: dos de los microtúbulos de cada triplete del cuerpo basal se extienden hasta esta región, que contiene, además de estos 9 dobletes periféricos, un par de microtúbulos centrales, que originan una estructura 9+2. A diferencia del par central, en los dobletes periféricos los microtúbulos comparten protofilamentos. Estos dobletes están conectados entre sí por nexina, y al par central mediante espinas radiales. Además, el microtúbulo A está asociado a dos brazos de dineína, responsable del movimiento.
- Zona de transición: se sitúa entre las dos anteriores. Contiene únicamente 9 dobles periféricos, conectando el corpúsculo basal con la membrana plasmática, mediante una placa basal.
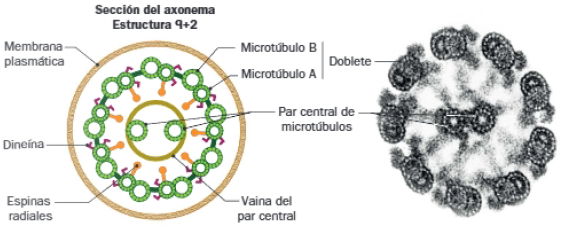
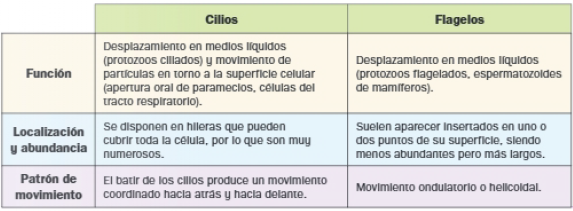
Comparación de cilios y flagelos
Ambos apéndices poseen estructura interna e igual mecanismo de producción de movimiento: la actividad motora de dineínas localizadas entre los microtúbulos del axonema, provoca el deslizamiento de unos dobletes sobre otros. Sus diferencias se resumen a continuación:
Huso mitótico
Hacia el final de la interfase, se duplica el centrosoma. El desplazamiento de los centrosomas hacia polos opuestos de la célula permite reorganizar el esqueleto de microtúbulos, desde ambos COMT hasta conformar el huso mitótico, constituido por 3 tipos de microtúbulos:
- Microtúbulos cinetocóricos.
- Microtúbulos del áster.
- Microtúbulos polares.
B. Filamentos intermedios
La denominación de los filamentos intermedios se debe a su grosor, que es intermedio al de microtúbulos y microfilamentos de actina.
Están formados por diferentes proteínas fibrosas, como queratinas en las células epiteliales, desmina en los sarcómeros o la proteína de neurofilamentos en las neuronas. Su ensamblaje lleva consigo enrollamientos helicoidales, que originan filamentos estables y apolares, semejantes a una cuerda.
El ensamblaje de filamentos intermedios comienza con la formación de dímeros de los polipéptidos que los conforman. Le sigue la formación de tetrámeros y protofilamentos.
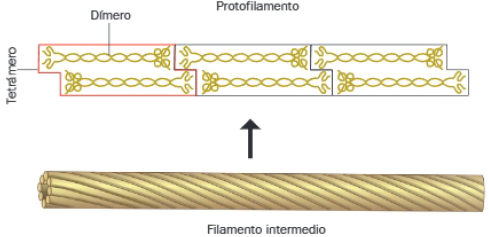
La unión lateral de protofilamentos origina un filamento de 10 nm (filamento intermedio).
Cabe puntualizar cuales son las funciones de los filamentos intermedios:
- Son muy abundantes en los animales, realizan funciones estructurales.
- Destaca la formación de armazón o red que se extiende desde la periferia nuclear hasta la membrana.
- Además de posicionar el núcleo, aporta estabilidad mecánica a la célula y elementos del citoesqueleto.
- En células epiteliales intervienen en la formación de uniones adhesivas.
- En el núcleo interfásico forman la lámina nuclear, por lo que su función también se relaciona con la división celular.
C. Microfilamentos de actina
Los microfilamentos de actina son los elementos del citoesqueleto más finos, con un diámetro de 7 nm, cuya estructura helicoidal deriva de la polimerización de la proteína citoesquelética más abundante, la actina.
Veamos algunas de sus características más importantes:
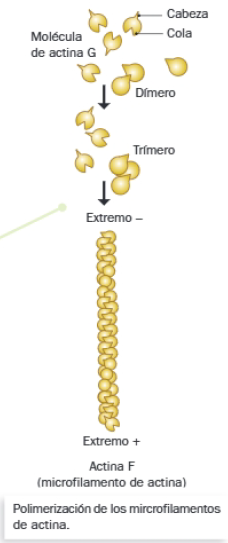
- Están formados por la unión de monómeros de actina G, moléculas individuales de esta proteína globular.
- En cada molécula de actina G se diferencian una cabeza puntiaguda y una cola protuberante. Su polimerización se basa en interacciones cabeza-cola entre monómeros diferentes, las cuales provocan un giro en cada monómero, dando lugar a la apariencia de doble hélice característica de los microfilamentos de actina o actina F (polimerizada).
- La polimerización de los microfilamentos consume ATP, y tiene lugar mediante adición de actina G a su extremo positivo. Dicha adición es reversible, por lo que, al igual que los microtúbulos, estos filamentos presentan un comportamiento dinámico.
Veamos a continuación las principales funciones de los microfilamentos de actina:
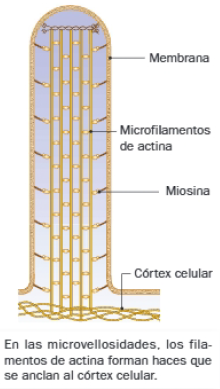
- Son especialmente abundantes bajo la membrana plasmática, a la que se asocian para formar el córtex celular; además de aportar soporte mecánico y determinar la forma celular, constituyen el origen de actuación de estos filamentos. Actúan en las interacciones de las células con la matriz extracelular y en interacciones entre células, concretamente en uniones adhesivas como los desmosomas.
- Forman el esqueleto mecánico de estereocilios y microvellosidades.
- Responsables de los movimientos celulares basados en proyecciones de membrana, tales como el movimiento ameboide y la emisión de pseudópodos.
- En los músculos esqueléticos y cardiaco, se asocian con miosina para formar unidades denominadas sarcómeros, responsables de la contracción muscular.
- Forman un anillo contráctil responsable de la citocinesis en las células animales.
- Sirven de vía para el transporte intracelular de orgánulos y vesículas.
ORGÁNULOS CENTRALES
La complejidad que caracteriza el citoplasma eucariota deriva de la variedad de orgánulos que presenta, la mayoría de los cuales están rodeados de membrana.
Los orgánulos membranosos compartimentan el citoplasma y las funciones celulares de eucariotas, permitiendo que se desarrollen eficazmente.
Algunos de estos orgánulos se encuentran estrechamente relacionados, como el retículo endoplasmático y el aparato de Golgi. Son orgánulos muy voluminosos, cuyas membranas, comunicadas mediante vesículas, ocupan buena parte del citoplasma, que conforma el denominado sistema de endomembranas.
Además del núcleo, en el citoplasma existen otras estructuras membranosas: los lisosomas, las vacuolas y los orgánulos energéticos (peroxisomas, mitocondrias y cloroplastos), a los que se les atribuye un origen endosimbiótico.
Ribosomas
Los ribosomas son orgánulos no membranosos encargados de dirigir el proceso de traducción o síntesis de las proteínas de la célula. Están integrados por dos subunidades, mayor y menor, compuestas por proteínas, con función estructural, y ARNr, que cataliza la formación de enlaces peptídicos (ribozima).
Las células eucariotas presentan dos tipos de ribosomas, que difieren en su coenficiente de sedimentación:
- Ribosomas 80S (formados por subunidades 60S y 40S): se encuentran libres en el citoplasma, asociados a las membranas del núcleo y del retículo endoplasmático rugoso.
- Ribosomas 70S (formados por subunidades 50S y 30S): son típicamente procariotas, por lo que se localizan en la matriz de mitocondrias y cloroplastos.
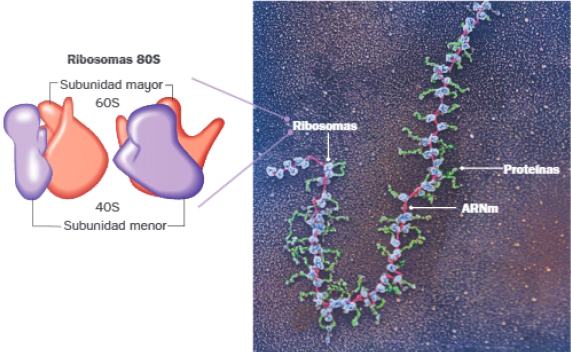
Los polisomas o polirribosomas están formados por un conjunto de ribosomas que traducen simultáneamente el mismo ARNm.
Pueden observarse en el citoplasma de procariotas y eucariotas, cuando una proteína debe sintetizarse adecuadamente.
Retículo endoplasmático
Está formado por una red de membranas interconectadas que se extiende por el citoplasma. Estas delimitan un espacio interno o lumen, en el que se desarrollan procesos de síntesis de biomoléculas. Se distinguen dos tipos, con diferente estructura y función: el retículo endoplasmático rugoso (RER) y el liso (REL).
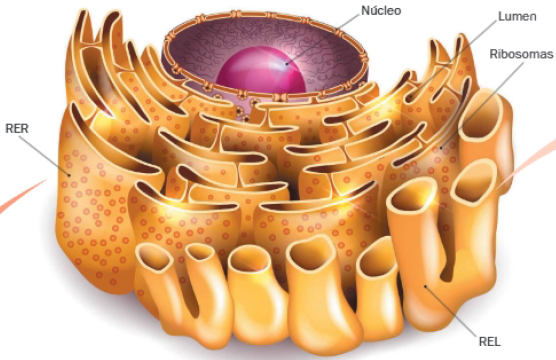
A. Retículo endoplasmático rugoso
El RER está formado por una red interconectada de sacos membranosos aplanados (cisternas), comunicados con la envoltura nuclear. Se denominación se debe a la presencia de ribosomas, adheridos a su cara citosólica.
El RER se relaciona con la producción, modificación y control del destino de las proteínas:
- Síntesis de proteínas en los ribosomas de la cara citosólica de sus membranas. Se translocan al retículo a medida que se traducen, para ser incorporadas a sus membranas o liberadas en el espacio luminal. Las primeras pasarán a formar parte de otras membranas, mientras que las segundas serán secretadas por la célula o irán a parar al interior del retículo, del aparato de Golgi o de lisosomas.
- Modificaciones postraduccionales de las proteínas sintetizadas, tales como plegamientos, ensamblaje de varias subunidades o glucosilaciones (formación de glucoproteínas).
- Procesos de control de las proteínas obtenidas. Se presentan alguna anomalía, salen del retículo para ser degradadas en el citoplasma.
- Empaquetamiento de las moléculas sintetizadas en vesículas de membrana para su transporte. Las proteínas que desempeñan su función en la membrana plasmática o en los lisosomas, así como las que deben ser secretadas por la célula, pasan al aparato de Golgi.
B. Retículo endoplasmático liso
El REL está formado por una red de túbulos membranosos, conectados entre sí y a las membranas del retículo rugoso. Recibe esta denominación porque, a diferencia del anterior, no se asocia con ribosomas.
Si bien el REL se encarga de la síntesis de la mayoría de los lípidos de la célula, también desempeña otras funciones:
- En su cara citoplasmática se sintetizan los glicerofosfolípidos de las membranas celulares.
- Sintetiza otro componente fundamental de las membranas, el colesterol, además de la molécula precursora de otros fosfolípidos y glucolípidos de membrana, la ceramida.
- Síntesis de hormonas esteroideas a partir de colesterol.
- En órganos como el hígado y el riñón, metaboliza tóxicos liposolubles (detoxificación), procedentes de medicamentos, drogas o células extrañas, y los transforma en compuestos liposolubles que se pueden eliminar a través de la orina.
- Libera glucosa de los gránulos de glucógeno adheridos a él en los hepatocitos.
- En células musculares se especializa en acumular calcio, cuya liberación al citoplasma provoca la contracción muscular, por lo que pasa a denominarse retículo sarcoplásmico.
- Empaquetamiento en vesículas de transporte de las moléculas sintetizadas, que se dirigen al aparato de Golgi.
Aparato de Golgi
Es un orgánulo que está formado por cisternas apiladas, que se relacionan con las demás membranas celulares mediante vesículas. Cada conjunto de sáculos apilados se denomina dictiosoma, y constituye la unidad funcional del aparato de Golgi. Los dictiosomas presentan polaridad, pues en ellos se diferencian la cara cis y cara trans.
Cara cis (convexa, orientada al núcleo): recibe las moléculas producidas en el retículo para comenzar su modificación. También se denomina cara de formación, pues en ella comienza la síntesis de las moléculas que se originan en el aparato de Golgi. Todas estas moléculas son sometidas a nuevas modificaciones a medida que las cisternas avanzan hacia la cara trans.
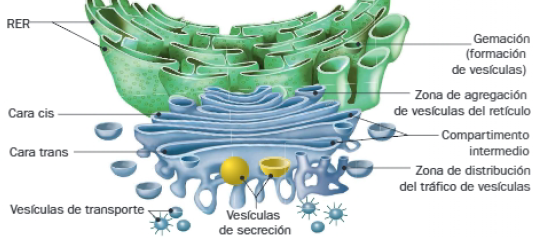
Cara trans (cóncava, orientada hacia el citoplasma) o cara de maduración: una vez finalizado el proceso de maduración, las moléculas alcanzan esta cara del dictiosoma. A continuación, son empaquetadas en vesículas de transporte antes de dirigirse a su destino.
Según la función que vayan a desempeñar las moléculas que transportan, las vesículas se fusionan con la membrana plasmática, para secretar su contenido mediante un proceso de exocitosis, o se dirigen a lugares de la célula para permanecer en ella (lisosomas, membrana plasmática, matriz extracelular o pared celular).
Tanto las vesículas que se forman a partir de las membranas del retículo como el aparato de Golgi lo hacen por gemación, y una vez que alcanzan la membrana diana, se fusionan con ella, y posibilitan la evacuación de su contenido.
Finalmente vamos a destacar las funciones principales del aparato de Golgi:
- Modificación y maduración de los lípidos procedentes del REL y de las proteínas procedentes del RER, hasta alcanzar su forma funcional. Alguna de estas modificaciones consiste en glucosilaciones, por lo que en el aparato de Golgi también se sintetizan glucoproteínas.
- Síntesis de glucolípidos y esfingomielina a partir de ceramida procedente del RER.
- Síntesis de hemicelulosa y pectinas de la pared celular vegetal, y de glucoaminoglucanos de las matrices extracelulares.
- Empaquetamiento y transporte de las moléculas anteriores, en el interior de vesículas.
- Génesis de lisosomas y de otras vesículas, como las que forman vacuolas vegetales o las que conforman el fragmoplasto durante la división en estas células.
Vesículas especializadas
En la célula existe vesículas que realizan funciones diferentes a las de secreción y transporte. Estos orgánulos membranosos pueden proceder de vesículas del aparato de Golgi (lisosomas y vacuolas) o del retículo endoplasmático (peroxisomas).
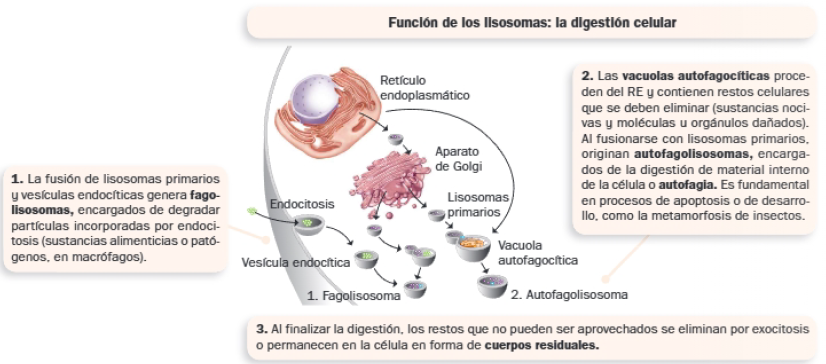
A. Lisosomas
Los lisosomas contienen enzimas hidrolíticas que catalizan reacciones de degradación de biomoléculas, por lo que se encargan de la digestión celular. La mayoría de las enzimas son hidrolasas ácidas: por ello, el interior de los lisosomas es ácido, y se debe a la presencia en su membrana de una bomba que introduce protones activamente. Se distinguen dos tipos de lisosomas:
- Lisosomas primarios: vesículas procedentes del aparato de Golgi que contienen enzimas.
- Lisosomas secundarios: formados por la fusión de los anteriores con otras vesículas, que originan vacuolas digestivas. Según la procedencia del material para degradar, reciben la denominación de fagolisosomas o autofagolisosomas.
B. Peroxisomas
Son pequeñas vesículas que contienen enzimas oxidativas (oxidasas) e intervienen en el metabolismo energético de la célula. Las reacciones oxidativas que llevan a cabo pueden generar peróxido de hidrógeno, nocivo para la célula; por ello, contienen una enzima que lo descompone, la catalasa.
En cuanto a sus funciones principales podemos destacar las siguientes:
- Obtención de energía mediante la oxidación de ácidos grasos (almacenados en las semillas).
- Este proceso ocurre en los vegetales, como al igual ocurre el mismo proceso en los animales en las mitocondrias.
- Intervienen en procesos de detoxificación, fundamentales para el hígado y el riñón (fotorrespiración).
C. Vacuolas
A pesar de que se diferencian varios tipos de vacuolas, todas tienen en común su elevado contenido hídrico. Los protozoos pueden presentar vacuolas pulsátiles, encargadas de evacuar agua que entra en la célula por ósmosis. Sin embargo, entre ellas destacan las vacuolas de células vegetales, que pueden fusionarse hasta originar una gran vacuola.
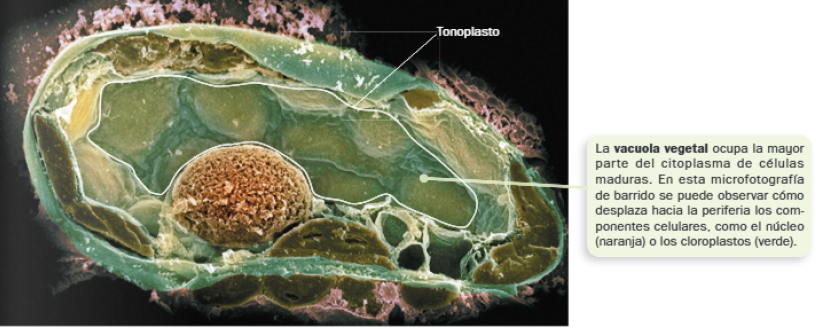
Cabe destacar las principales funciones de las vacuolas vegetales:
- Mantenimiento de la turgencia celular (principal), responsable del crecimiento de estas células por medio de un económico mecanismo.
- La acumulación de solutos en su interior provoca la entrada de agua por ósmosis y el incremento de la presión osmótica sobre su membrana (tonoplasto).
- El aumento del volumen de la vacuola desplaza los demás componentes citoplasmáticos, que ejercen presión sobre la pared celular (presión de turgencia).
- Determinadas vacuolas pueden hacer la función de almacén de sustancias o de digestión celular a través de hidrolasas ácidas.
Orgánulos energéticos
Igual que los peroxisomas, las mitocondrias y los cloroplastos llevan a cabo reacciones que aportan energía a la célula. A diferencia de los primeros, producen energía en forma de ATP.
Las mitocondrias y los cloroplastos son los únicos orgánulos de membrana doble, lo cual se atribuye a su origen endosimbiótico. La disposición de sus membranas genera compartimentos funcionales en estos orgánulos.
A. Mitocondrias
En el interior de las mitocondrias se diferencian dos compartimentos, el espacio entre sus dos membranas y una matriz interna, generados por la disposición de sus componentes:
- Membrana mitocondrial externa: límite externo altamente permeable por la presencia de porinas, canales de permiten el paso de moléculas de pequeño tamaño.
- Espacio intermembrana o perimitocondrial: localizado entre ambas membranas y ocupado por una matriz de composición similar al citosol.
- Membrana mitocondrial interna: es menos permeable que la anterior y se caracteriza por presentar numerosas invaginaciones perpendiculares al eje mayor del orgánulo, denominadas crestas mitocondriales. En ellas se localizan las cadenas de transporte de electrones y las ATP sintasas, complejos enzimáticos localizados en la cara matricial de las crestas.
- Matriz mitocondrial: este compartimento se corresponde con el espacio interno de la mitocondria, y en su composición destaca la presencia de varias copias de ADN mitocondrial, ribosomas 70S, ARN mitocondrial y diversas enzimas (las responsables de la expresión de su genoma y de las principales reacciones del metabolismo oxidativo).
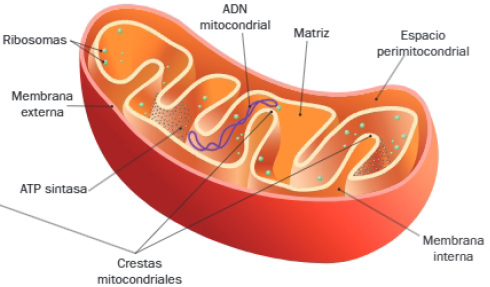
Finalmente destacaremos las funciones principales de las mitocondrias:
- Son los principales centros de producción de energía por respiración celular.
- En la matriz mitocondrial, además de la síntesis de proteínas mitocondriales, tienen lugar los siguientes procesos metabólicos:
- Beta oxidación de los ácidos grasos.
- Descarboxilación oxidativa del ácido pirúvico procedente de la glucólisis.
- Ciclo de Krebs.
- En la membrana mitocondrial interna se localiza la cadena respiratoria o transportadora de electrones. Este proceso genera un gradiente electroquímico y está acoplado a la síntesis de ATP en las ATP sintasas localizadas en esta membrana, proceso conocido como fosforilación oxidativa.
B. Cloroplastos
Los cloroplastos pertenecen a una familia de orgánulos denominados plastos o plásticos, que presentan el mismo genoma y doble membrana, pero diferente estructura y función. a pesar de que existen plástidos sin pigmentos, los cloroplastos pertenecen al grupo de los plastos pigmentados, pues contienen pigmentos fotosintéticos entre los que destacan las clorofilas, responsables de su coloración verde y de la absorción de luz.
Constan de los siguientes componentes:
- Membrana plastidial externa e interna: están separadas por el espacio intermembranoso. Todas ellas presentan características semejantes a las mitocondriales.
- Estroma: espacio situado entre la membrana plastidial y la tilacoidea. Es análogo a la matriz mitocondrial, por lo que además de varias moléculas de ADN, enzimas relacionadas con su metabolismo y ribosomas 70S, contiene diversas inclusiones, como gránulos de almidón, y enzimas implicadas en la fijación del carbono, como la RuBisCo.
- Tilacoides: conjunto de sacos membranosos interconectados, paralelos al eje mayor. Pueden estar apilados, formando conjuntos denominados grana, los cuales se conectan por medio de tilacoides que atraviesas el estroma (tilacoides intergrana). Estas membranas, además de las cadenas de transporte electrónico y los complejos ATP sintasa, contienen pigmentos fotosintéticos en los fotosistemas que captan la luz solar. Las membranas de los tilacoides delimitan un tercer compartimento, el espacio tilacoidal.
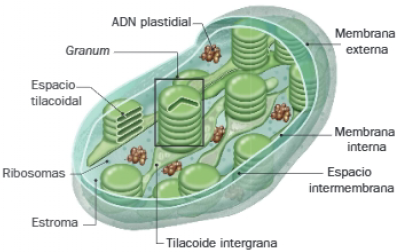
Finalmente destacaremos las funciones principales de los cloroplastos:
- Los cloroplastos llevan a cabo funciones de fotosíntesis oxigénica, que permite a los organismos fotosintéticos transformar biomoléculas inorgánicas en orgánicas. La fotosíntesis se desarrolla en dos etapas:
- Fase Lumínica: conjunto de reacciones que ocurren en las membranas tilacoidales y son dependientes de luz, por lo que en ellas es fundamental el papel de la clorofila. Igual que ocurre en la membrana mitocondrial interna, en esta fase hay transporte electrónico acoplado a la síntesis de ATP.
- Fase oscura: se desarrolla en el estroma y no es dependiente de luz solar.
- Además de la síntesis de proteínas y lípidos de membrana plastidiales, en el estroma de los cloroplastos tienen lugar reacciones del metabolismo del nitrógeno, concretamente la asimilación de nitratos a sulfatos y la reducción de nitrito a amoníaco.
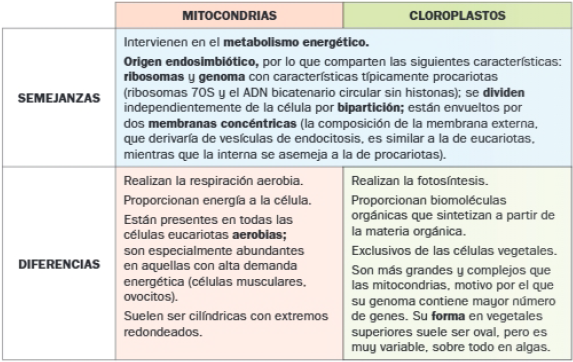
C. Semejanzas y diferencias entre mitocondrias y cloroplastos
Fuente: Biología 2 (Mc Graw Hill).