
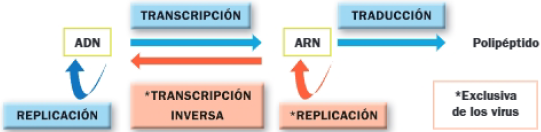
1. DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR
El dogma central de la biología molecular describe el flujo de la información genética: la transmisión del mensaje genético de una generación a la siguiente, y la expresión a su forma funcional, las proteínas.
Pese a que las características que definen la ciencia son contrarias a las que definen los dogmas, en este caso particular se utiliza dicha denominación para destacar la importancia de esta teoría.
El conocimiento actual sobre el flujo de la información deriva de un conjunto de contribuciones realizadas a lo largo del siglo XX. Entre ellas destacan las siguientes:
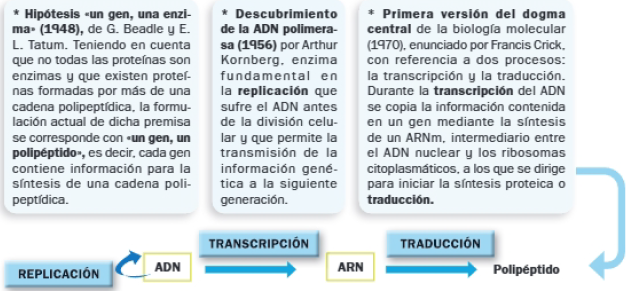
El conocimiento del ciclo replicativo de los virus llevó a modificar el dogma inicial:
- Los virus pueden tener ADN o ARN como material genético. La enzima ARN replicasa se encarga de replicar el ARN vírico.
- Los retrovirus, virus con ARN como material genético, poseen la enzima transcriptasa inversa o retrotranscriptasa, capaz de sintetizar ADN tomando como molde una cadena de ARN. Sintetizan ADN a partir de dicho ARN durante el proceso denominado transcripción inversa.
2. EL ADN CONTIENE LA INFORMACIÓN GENÉTICA
Las características químicas del ADN hacen que sea la molécula idónea para portar la información biológica de los seres vivos.
La molécula de ADN contiene la información genética codificada en la secuencia de bases nitrogenadas que conforman cada gen. Sin embargo, y pese a conocerse la existencia de esta desde 1869 gracias a Friedrich Miescher, antes de corroborar experimentalmente este hecho se consideraba que eran las proteínas las portadoras del mensaje genético.
Antes de comprobar que se trataba del ADN, se había deducido que la molécula encargada de almacenar la información genética debía cumplir una serie de requisitos.
Aquí veremos a continuación las características de la molécula portadora de la información genética:
- Es transmisible, es decir, la información que contiene pasa de una generación a la siguiente.
- Presenta capacidad de autorreplicación, por tanto, puede originar copias de sí misma, posibilitando el reparto equitativo entre las células hijas durante la división celular y el consiguiente mantenimiento de la identidad de una misma estirpe celular.
- Es químicamente estable, por lo que no sufre alteraciones bruscas, permitiendo una transmisión de información fiable generación tras generación.
- Es susceptible de sufrir pequeños cambios químicos: su frecuencia de mutación aporta cierta variabilidad genética, que favorece la actuación de la selección natural y explica los fenómenos evolutivos.
¿Cómo se logró la identificación de la molécula portadora de la información genética?
La atribución de la función portadora del mensaje genético al ADN fue posible gracias a una serie de experimentos desarrollados durante la primera mitad del siglo XX.
Entre ellos se encuentran los experimentos de Frederick Griffith (1928). Estos mostraron que cepas no virulentas del Streptococcus pneumoniae se transformaran en virulentas al entrar en contacto con el contenido celular de aquellas que sí causaban neumonía, de lo que pudo deducir que en su interior se encontraba la molécula causante de dicha información, a la que denominó «factor transformante».
La identificación del principio transformante de Griffith llegó en 1944 de la mano de Oswald Avery y colaboradores, pues observaron que la capacidad transformante de las cepas virulentas de Streptococcus pneumoniae desaparecía al añadir enzimas que degradan el ADN.
Posteriormente, numerosas investigaciones, como las desarrolladas por Martha Chase y Alfred Hershey en 1952, apoyaron este indicio. El uso de virus bacteriófagos marcados radiactivamente les permitió concluir definitivamente que la molécula que contiene la información genética es el ADN, en lugar de las proteínas.
A. El experimento de Frederick Griffith
Metodología y resultados
En búsqueda de una vacuna para la neumonía, Griffith realizó experimentos con dos cepas de la bacteria Streptococcus pneumoniae:
- Cepa R: bacterias de superficie rugosa, sin cápsula e inocuas.
- Cepa S: bacterias de superficie lisa, con cápsula y virulentas.
Al inocular diferentes muestras de ambas en ratones, obtuvo los siguientes resultados:
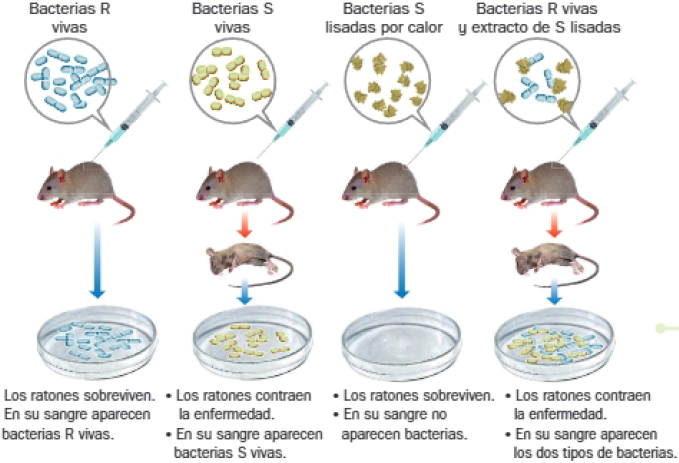
Conclusiones:
- La información responsable de la formación de la cápsula (que proporciona virulencia) está presente en el contenido de las bacterias S y se transfiere de las bacterias S a las R.
- Este componente es un portador activo de la información genética, pues se mantiene activo a pesar de la destrucción de las células que lo contienen.
B. Experimento de Martha Chase y Alfred Hershey
Metodología y resultados
Los bacteriófagos, virus que infectan bacterias, poseen una cápside proteica (cabeza) que contiene el ADN en su interior. Para iniciar su multiplicación, inyectan su ADN en el interior de una bacteria, mientras que la cápside permanece fuera de la célula.
Marcaron radiactivamente los componentes de bacteriófago T2; prepararon dos cultivos: uno contenía el isótopo 32p, que pasaría a formar parte del ADN del virus; el otro cultivo contenía el isótopo 35s que se incorporaría a las proteínas de la cápside. Infectaron bacterias con sendos grupos de virus y midieron su radiactividad.
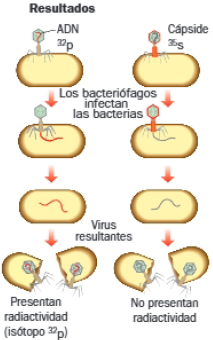
Conclusiones:
Observaron que solo aparecía radiactividad en el interior de las bacterias y en los virus resultantes de la multiplicación en el cultivo con virus marcados con 32p. El 35s no entraba en las bacterias, se quedaba en la superficie. Concluyeron que el material genético es el ADN y no las proteínas.
3. CARACTERÍSTICAS DEL GENOMA PROCARIOTA Y EUCARIOTA
El término genoma alude al conjunto de genes del organismo. Como en mucho otros aspectos, existen importantes diferencias entre el genoma de los dos grandes tipos de células.
Las diferencias entre los genomas procariotas y eucariotas están relacionadas con la estructura y cantidad de ADN, y el modo en que en él se organiza la información genética.
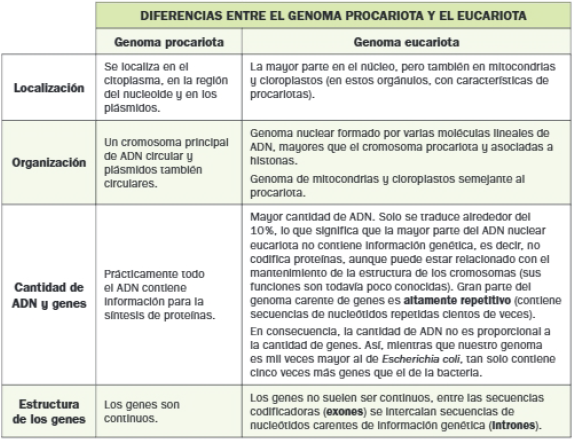
A diferencia de lo que ocurre entre el número de genes y la complejidad de los seres vivos eucariotas, la cantidad y longitud de los intrones sí está directamente relacionada con la complejidad de estos organismos, pues los exones suponen una ventaja evolutiva: además de facilitar la recombinación meiótica al aumentar la distancia entre los exones, las proteínas codificadas por varios exones pueden presentar pequeñas variantes cada vez que se traducen. Todo ello aporta variabilidad genética, la cual favorece la actuación de la selección natural.
4. LA REPLICACIÓN DEL ADN
La replicación tiene como finalidad duplicar el ADN para dotar a la célula de dos copias de su genoma y posibilitar el reparto equitativo de material genético entre las células hijas.
Su importancia radica en que permite el mantenimiento de la identidad celular, ya que la síntesis de moléculas prácticamente idénticas a la inicial asegura que se mantenga el mensaje genético, es decir, que sea transmitido fielmente a la siguiente generación.
Durante la fase S del ciclo celular, antes de la división de la célula, se sintetiza una copia de cada molécula de ADN. Mientras que en procariotas tiene lugar en el citoplasma, en eucariotas ocurre en el interior del núcleo.
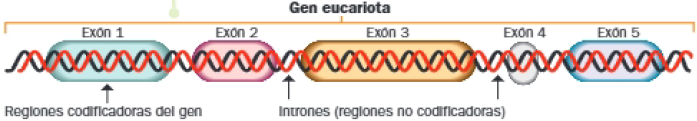
Un proceso semiconservativo
La replicación es semiconservativa, lo que significa que las moléculas de ADN resultantes conservan una cadena original, unida a otra complementaria sintetizada en este proceso (cadena de nueva síntesis).
Esto es posible gracias a la separación de las dos cadenas originales (hebras molde) al comienzo de la replicación, pues permite la actuación de diversas enzimas y la síntesis de sus hebras complementarias.
Antes de la demostración experimental de este hecho, que tuvo lugar en 1957 gracias a Matthew Meselson y Franklin Stahl, existían varias hipótesis sobre el mecanismo de replicación. Entre las alternativas a la semiconservativa, propuesta por Watson y Crick en 1953 con el modelo de doble hélice, se encontraban las hipótesis conservativa y dispersiva.
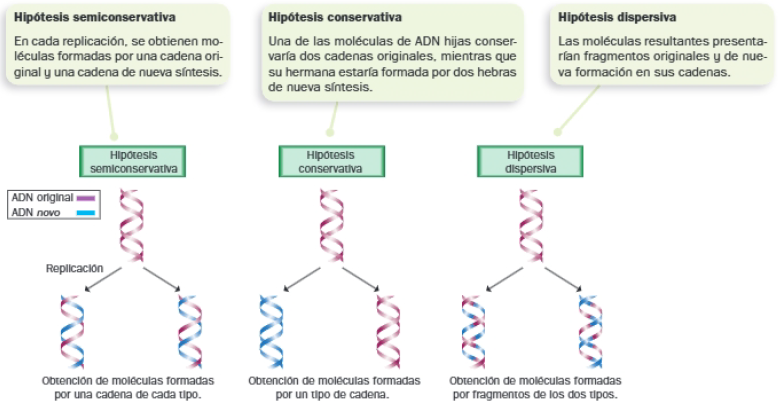
A. El experimento de Meselson y Stahl
Metodología y resultados
Para tratar de averiguar qué hipótesis sobre la replicación del ADN era la correcta, prepararon cultivos de Escherichia coli en medios nutritivos con diferentes fuentes de nitrógeno: uno de ellos contenía el isótopo pesado del nitrógeno, el otro contenía el isótopo habitual en el ADN, más ligero.
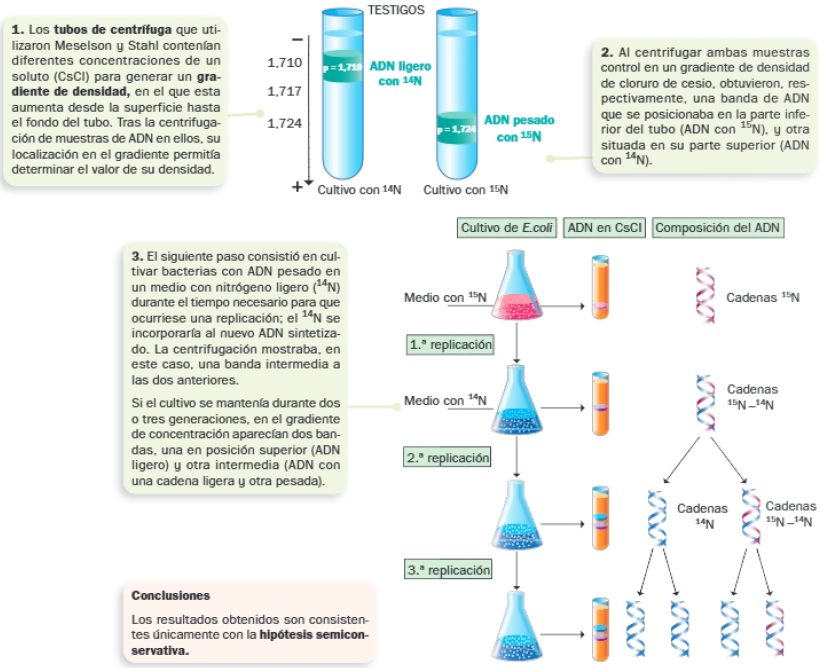
Etapas de la replicación en procariotas
El mecanismo fundamental de la replicación del ADN es común a procariotas y eucariotas. Este proceso ocurre en las siguientes fases, gracias a la actuación de diversas enzimas:
- Iniciación: apertura de la doble hélice, que permite el acceso a las cadenas simples.
- Elongación: síntesis de cadenas complementarias a las hebras originales, que actúan como molde, y corrección de errores.
- Terminación: las moléculas resultantes adquieren la configuración helicoidal.
A. Inicio de la replicación
La replicación se inicia en secuencias concretas del ADN, denominadas orígenes de replicación. A partir de ellas, se inicia la actuación de las siguientes enzimas:
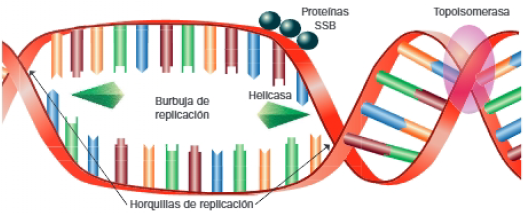
- Helicasas: rompen los enlaces de hidrógeno que mantienen unida la doble hélice, y producen la desespiralización de la molécula y la separación de sus cadenas.
- Topoisomerasas: evitan el superenrollamiento que se generaría por la separación de la doble hélice, para lo que realizan cortes y uniones en determinados puntos de esta.
- Proteínas SSB: estabilizan las cadenas separadas, y evitan que se vuelvan a unir.
Como consecuencia, se forma la burbuja de replicación, regiones de los cromosomas donde la doble hélice se encuentra separadas y accesible a las enzimas que dirigen la síntesis de ADN. Se corresponden, por tanto, con los lugares de replicación, cuyos extremos se denominan horquillas de replicación, generadas por la acción de las helicasas.
B. Elongación de las nuevas cadenas de ADN
El papel de las enzimas ADN polimerasas es fundamental en la replicación del ADN, especialmente el de la ADN polimerasa III, encargada de unir los nucleótidos complementarios a las cadenas molde.
Características de la ADN polimerasa III
- Recorre las hebras molde de ADN en sentido 3–5, y sintetiza la cadena complementaria y antiparalela, por lo que une nucleótidos en sentido 5–3.
- Es incapaz de iniciar la síntesis, ya que necesita un pequeño fragmento de ARN, denominado cebador o primer, a cuyo extremo 3 libre añade nucleótidos monofosfato.
- Debido a que las cadenas de ADN son antiparalelas y que esta enzima las recorre en un único sentido, la síntesis de una de ellas es más lenta y fragmentada (síntesis discontinua, mediante los denominados fragmentos de Okazaki).
- Usa nucleótidos trifosfato, a partir de los que obtiene, por hidrólisis del enlace fosfodiéster, los nucleótidos monofosfato de A, G, T y C presentes en el ADN y energía para su unión.
Diferencias en la replicación de eucariotas
La replicación del ADN fue estudiada por primera vez en Esscherichia coli. Más tarde se comprobó que las diferencias que existen en el proceso eucariota no afectan a su mecanismo, y que derivan de la mayor complejidad del genoma eucariota.
Principales diferencias en la replicación de eucariotas
- El ADN eucariota está asociado a las histonas formando la cromatina, por lo que la replicación del ADN va a compañada de la necesaria síntesis de las histonas.
- Existen 5 ADN polimerasas, frente a las 3 de las procariotas. Una de ellas interviene en la replicación del ADN mitocondrial.
- Debido que las eucariotas presentan mayores cantidades de ADN, para acortar la duración del proceso, existen múltiples orígenes de replicación. Por ello, existen numerosas unidades de replicación en su genoma, denominadas replicones.
- La velocidad de replicación es menor en eucariotas.
- Los fragmentos de Okazaki son más cortos en eucariotas (100-200 nucleótidos frente 1000-2000 en procariotas).
5. LA TRANSCRIPCIÓN
La transcripción es la síntesis de ARN a partir de información contenida en el ADN.
La transcripción génica implica dos procesos fundamentales: la transcripción y la traducción.
Si en la transcripción se sintetiza un ARN mensajero, irá seguida de la traducción de este para originar una proteína.
Características de la transcripción
Las características fundamentales de este proceso son las siguientes:
- Tiene lugar durante la interfase del ciclo celular, sobre todo, en los periodos G1 y G2.
- En las procariotas ocurre en el citoplasma; en las células eucariotas, fundamentalmente en el núcleo, pero también en la matriz de mitocondrias y en el estroma de los cloroplastos.
- Se transcribe una de las dos cadenas de ADN, la denominada cadena molde, o no codificante, que tiene la secuencia complementaria de la ARN transcrito; en este caso, la guanina es complementaria a la citosina y el uracilo de la adenina.
- La cadena que no transcribe se denomina cadena codificante y tiene la misma secuencia que el ARN transcrito, con la excepción de que, en vez de timina, presenta uracilo.
- Durante la transcripción, las ARN polimerasas leen la cadena molde en sentido 3–>5 y sintetizan el ARN en sentido 5–>3.
Etapas de la transcripción
Desde el punto de vista molecular, podemos difinir un gen como una unidad de transcripción, es decir, una región del genoma con la información necesaria para la síntesis de ARN.
Al igual que la replicación, tanto en eucariotas como en procariotas, la transcripción conste de 3 etapas: Inicio, elongación y terminación.
A continuación analizaremos estas 3 etapas en procariotas, que han servido de base para comprender los mecanismos más complejos en las eucariotas:
A. Inicio de la transcripción
Para que dé inicio la transcripción, la célula procariota necesita:
- Una ARN polimerasa ADN dependiente, también llamada transcriptasa: la enzima responsable de la transcripción es una polimerasa que sintetiza ARN tomando como molde una cadena de ADN.
- Factor sigma: polipéptido independiente, subunidad necesaria para la unión de la ARN polimerasa al promotor.
El inicio se produce del siguiente modo:
- Unión del factor sigma al promotor.
- Apertura de la burbuja de transcripción, al separarse de las dos cadenas del ADN en esa región. Intervienen enzimas helicasas.
- Unión de la ARN polimerasa.
- Liberación del factor sigma.
B. Elongación del ARN
La formación de una cadena de ARN requiere:
- Una cadena molde de ADN.
- ARN polimerasa.
- Nucleótidos trifosfato de A, U, C y G.
La ARN polimerasa lee la cadena en sentido 3–>5 y una nucleótidos monofosfato en sentido 5–>3 mediante enlaces fosfodiéster, introduciendo en la cadena de ARN en formación de nucleótidos complementarios de la cadena de molde.
Para ello, utiliza nucleótidos trifosfato, rompiendo el enlace entre los grupos fosfato primero y segundo; de este modo, obtiene nucleótidos monofosfato y energía para la unión entre ellos.
C. Terminación de la transcripción
La ARN polimerasa se detiene cuando encuentra la secuencia terminadora, que le indica el final de la transcripción.
El ARN queda libre, la ARN polimerasa se separa del ADN, la horquilla de transcripción se cierra y el ADN recupera su forma original.
El ARN transcrito puede ser:
- Un ARN mensajero funcional, que ya puede ser traducido a la par que se transcribe.
- Un ARN ribosómico o ARN transferente; en este caso, deben sufrir modificaciones para ser funcionales.
6. LA TRADUCCIÓN
La traducción es el proceso mediante el cual las instrucciones del ADN, transcritas en forma de ARNm, son interpretadas y transformadas en la secuencia de aminoácidos, y originan un polipéptido o una proteína.
El ARNm contiene la información para la síntesis de una cadena polipeptídica o una proteína
El código genético
Puede definirse como la relación entre la secuencia de nucleótidos del ARNm y la secuencia de aminoácidos de un polipéptido o proteína.
A. Características del código genético
A partir de los años 50 del siglo XX, diversas experiencias contribuyeron a perfilar las características del código genético.
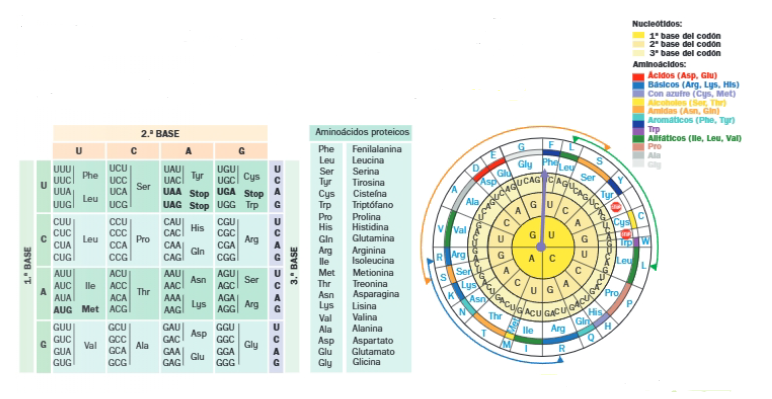
La secuencia de bases del ARNm se lee de 3 en 3. 3 bases del ARNm forman un triplete o codón que es la señal para un aminoácido. El número mínimo de bases para codificar un aminoácido se deduce fácilmente, teniendo en cuenta que hay 20 aminoácidos proteicos y solo 4 bases en el ARNm (A, G, C y U).

Hay más codones que aminoácidos codificados: 64 codones diferentes para tan solo 20 aminoácidos proteicos; por lo tanto, hay codones que codifican para el mismo aminoácido y se dice que el código presenta degeneración.
Los codones sinónimos son aquellos que codifican para el mismo aminoácido. Muchos de los codones sinónimos se diferencian entre sí en la tercera base del triplete. Una consecuencia de la degeneración es que disminuye la posibilidad de que una mutación puntual (cambio de una base en el ADN) provoque el cambio de un aminoácido en la proteína.
No presenta imperfecciones porque cada codón codifica para un único aminoácido.
Cada nucleótido forma parte de un único codón; no hay solapamientos entre los tripletes ni espacios entre ellos. Los codones se leen de forma ordenada y continua, sin interrupciones, desde el punto de inicio de la lectura hasta el final.
El codón de inicio es AUG. Los codones de terminación son aquellos que no codifican para ningún aminoácido; estos son: UAA, UAG y UGA; se conocen como codones sin sentido.
Todos los seres vivos utilizan el mismo código genético, salvo alguna excepción puntual en algún codón (en mitocondrias humanas y de otros mamíferos, y en determinadas bacterias).
B. El descifrado del código genético
Descifrar el código genético supuso asignar cada codón a su aminoácido correspondiente.
Para ello fue fundamental la enzima denominada polinucleótido fosforilasa, cuyo descubrimiento en 1955 le valió a Severo Ochoa el Premio Nobel. La polinucleótido fosforilasa permite sintetizar in vitro cadenas de ARN con una composición determinada, sin necesidad de un molde de ADN. Tan solo necesita un cebador, nucleótidos difosfato y magnesio.
En primer lugar, se sintetizaron ARNm con un único tipo de nucleótido: poli-U, poli-A, poli-C y poli-G y se vio que polipéptido se originaba al traducirlos.
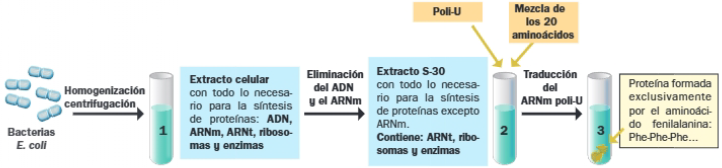
Para descifrar el resto de codones, se recurrió a ARNm sintético con secuencia repetidas 2, 3 y 4 veces.
Repitiendo la experiencia con diferentes ARNm, se logró descifrar por completo el código genético, que se muestra a continuación en las dos formas en que suele representarse:
El gen ocupa un lugar específico, locus, en un cromosoma y contiene, además de la secuencia que se transcribe en ARN, los elementos necesarios para regular su transcripción.
Si el ARN sintetizado es un ARN mensajero, la traducción posterior originará un polipéptido o una proteína.
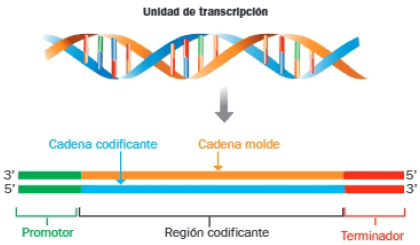
En la unidad de transcripción diferenciamos:
- Promotor: secuencia de nucleótidos que no se transcribe y marca el inicio. En procariotas y en las mitocondrias, es frecuente que un mismo promotor sirva para varios genes contiguos que se transcriben juntos. En el genoma nuclear de eucariotas, cada gen posee su propio promotor.
- Región codificante: contigua al promotor, formada por la secuencia de nucleótidos que se transcribe.
- Terminador: secuencia que indica el final de la transcripción (rica en G y C).
Etapas del proceso de traducción en procariotas
En la traducción diferenciamos las mismas tres fases que en la replicación y la transcripción: inicio, elongación y terminación, pero es necesaria una fase previa: la activación de los aminoácidos.
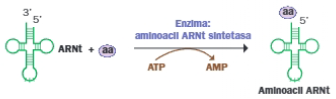
A. Activación de los aminoácidos
La activación consiste en la unión de cada aminoácido al ARN transferente que le corresponde, en el extremo 3. El proceso tiene lugar en el hialoplasma y necesita energía, aportada por el ATP.
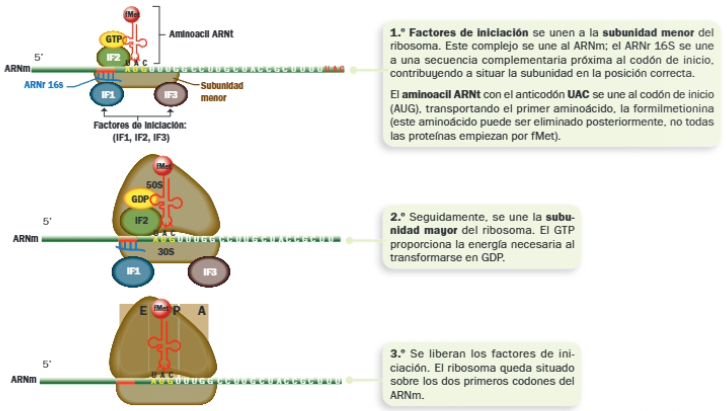
B. Inicio de la traducción
El inicio de la traducción requiere:
- Un ARNm.
- Un ribosoma.
- Aminoacil ARNt con el anticodón complementario del codón de inicio (de AUG). Este transporta el aminoácido formilmetionina (fMet) en procariotas.
- Factores de iniciación, que se unen a la subunidad menor del ribosoma y al aminoacil ARNt, facilitando la unión de ambos.
- GTP, que proporciona la energía necesaria.
El proceso se inicia del siguiente modo:
C. Elongación de la proteína
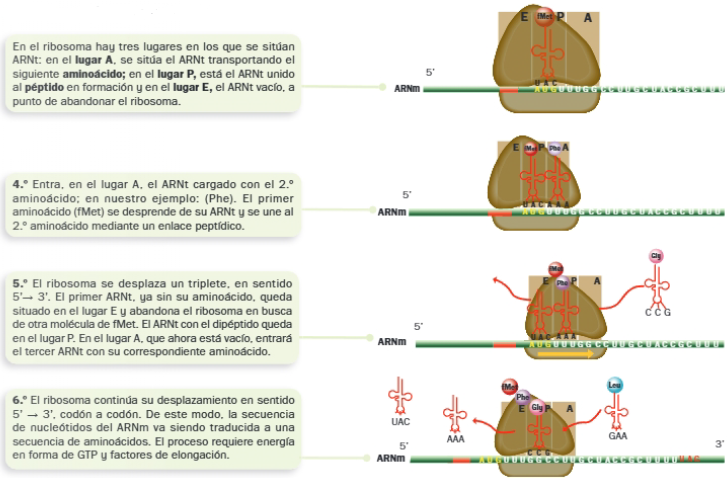
D. Terminación de la traducción
El proceso finaliza cuando el ribosoma se encuentra con uno de los tres codones sin sentido o codones de stop. Entonces, en el lugar A entra un factor de terminación. El péptido se desprende y el ribosoma se disocia en sus dos subunidades. El resultado es un péptido o una proteína:

Diferencias en la traducción de eucariotas
Las principales diferencias en relación con la traducción entre eucariotas y procariotas son las siguientes:
- En procariotas, transcripción y traducción pueden ser simultáneas: mientras está siendo ambos procesos tienen lugar en el citoplasma celular y, además, el ARNm de procariotas no necesita un proceso de maduración previo a la traducción. Esto es así también en mitocondrias y cloroplastos de células eucariotas. La transcripción del ADN nuclear eucariota tiene lugar en el núcleo, mientras que la traducción de los ARNm resultantes ocurre en el citoplasma, por lo tanto, son procesos separados espacial y temporalmente.
- En eucariotas la caperuza del extremo 5 del ARNm es necesaria al inicio de la transcripción, para la unión de la subunidad menor del ribosoma. El primer aminoácido es la metionina y se necesitan más factores de traducción que en procariotas.
7. REGULACIÓN DE LA EXPRESIÓN GÉNICA
La diferenciación celular es el proceso por el cual una célula, mediante la regulación de la expresión génica, se especializa y adquiere una morfología y una función determinada.
En los organismos pluricelulares, la regulación de la expresión génica está en el origen de la diferenciación celular.
A. Niveles de regulación de la expresión génica en eucariotas
La regulación de la expresión génica es notablemente más compleja que en procariotas.
Regulación pretranscripcional: Para que pueda transcribirse, el ADN debe ser accesible a las enzimas implicadas en el proceso, para ello es necesario que la cromatina se encuentre descondensada. El grado de compactación de la cromatina actúa controlando qué genes se transcriben y cuales no.
Regulación epigenética: la epigenética engloba el conjunto de cambios del ADN y/o las histonas, que no implican un cambio en la secuencia de bases, pero sí cambian el nivel de activación o inactivación de los genes.
Algunas modificaciones epigenéticas se pueden transmitir a las células hijas durante la división celular y de una generación a la siguiente. El conjunto de todos los cambios epigenéticos en un genoma se conoce como epigenoma.
Control de transcripción: la expresión de la mayoría de los genes eucariotas se controla al inicio de la transcripción, mediante los promotores y los factores de transcripción (TF).
Los promotores son regiones del ADN situados al inicio de un gen, regula la transcripción y constituyen la región reguladora del gen.
Los factores de transcripción son diversos y se unen a los promotores, favoreciendo o dificultando la actividad de la ARN polimerasa, según se necesite.
Controles postranscripcionales: el control de la maduración del ARN (la velocidad a la que se realiza y los procesos de splicing alternativo), el control del transporte del ARNm desde el núcleo hasta el citoplasma; el control de la degradación del ARNm (de su tiempo de permanencia en la célula dependerá cuantas veces puede ser traducido y, por lo tanto, la cantidad de proteína sintetizada).
Control de traducción: a través de los factores de iniciación.
Control del procesamiento de las proteínas: numerosas proteínas necesitan de modificaciones postraduccionales para ser funcionales.
Fuente: Biología 2 (Mc Graw Hill).