1 CATABOLISMO
En el catabolismo se produce la degradación (oxidación) de moléculas con el fin de obtener energía en forma de ATP. Durante las oxidaciones, se obtiene, además, coenzimas reducidas, que son necesarias para el anabolismo.
El catabolismo está conformado por una serie de rutas metabólicas convergentes: todas las degradaciones convergen en el denominado ciclo de Krebs, que se desarrolla en la matriz mitocondrial y supone la degradación total de la materia orgánica a inorgánica, tal y como se muestra en el siguiente esquema general del catabolismo:
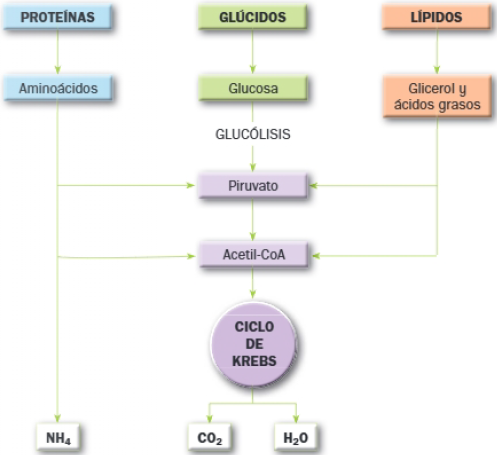
La principal fuente de energía de las células son los glúcidos; en concreto, la glucosa es el monosacárido que actúa como combustible universal; los lípidos como los triglicéridos también son degradados para obtener energía; en cambio, las proteínas cumplen otras funciones en las células y raramente se degradan los aminoácidos con fines energéticos.
Catabolismo de los glúcidos: fermentación y respiración aerobia
Los dos procesos fundamentales del catabolismo de los glúcidos son la fermentación y la respiración aerobia.
El siguiente esquema representa una visión global de los procesos implicados en el catabolismo de los glúcidos:
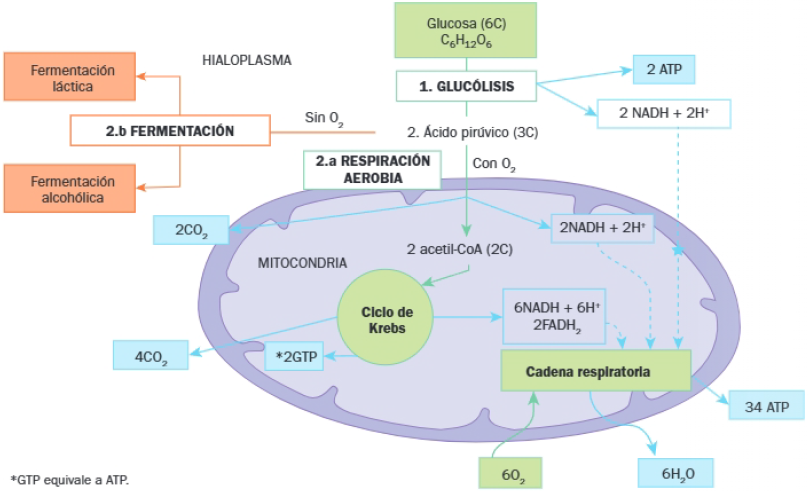
Se observa que tanto las fermentaciones como la respiración necesitan de un primer proceso, una ruta metabólica común en ambas: la glucólisis, que tiene lugar en el hialoplasma y en la que se obtiene ácido pirúvico o piruvato.
Destinos del piruvato:
- Si no hay oxígeno, el piruvato sigue una vía anaerobia: la fermentación, en el hialoplasma.
- Si hay oxígeno, el piruvato entre en una vía aerobia: la respiración aerobia, que tiene lugar en el interior de las mitocondrias.
El catabolismo en el hialoplasma tiene una serie de características:
- Se trata de degradaciones incompletas: el producto final es una molécula orgánica. Para extraer de ella toda la energía, el catabolismo debe continuar en las mitocondrias.
- Son reacciones anaerobia: no necesitan de oxígeno.
A. Glucólisis
La glucólisis es la degradación de la glucosa a piruvato (ácido pirúvico).
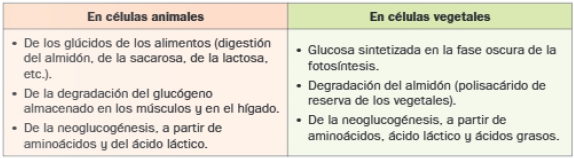
Origen de la glucosa:
Reacciones de la glucólisis:
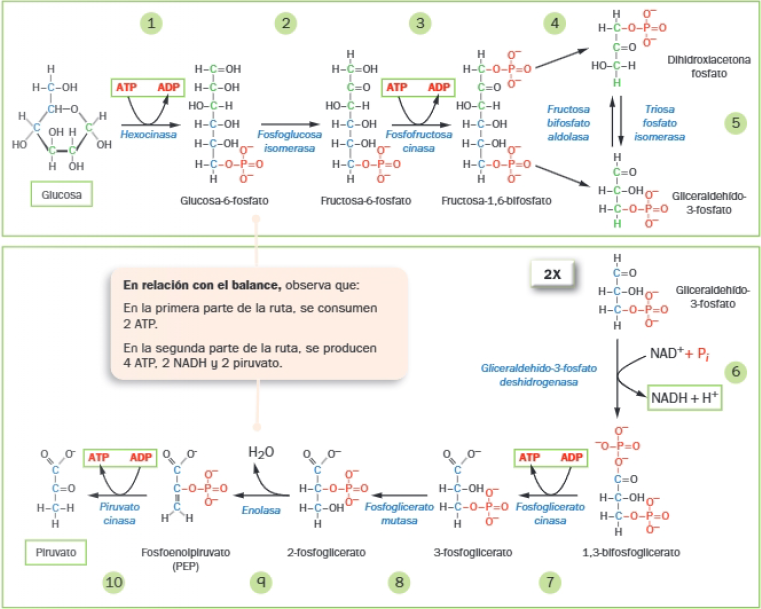
Las 10 reacciones de la glucólisis pueden agruparse en dos etapas:
- La primera parte concluye con el desdoblamiento de la fructosa 1,6-difosfato (hexosa) en dos triosas: gliceraldehído-3-fosfato y la dihidroxlacetona-fosfato.
- En la segunda parte, ambas triosas acaban convertidas en piruvato. Por eso, la segunda parte se produce dos veces (una por cada triosa).
Reacción global de la glucólisis:

La glucólisis cumple con los tres siguientes objetivos fundamentales:
- Obtener energía en forma de ATP.
- Obtener poder reductor en forma de NADH + H+.
- Obtener precursores metabólicos.
La glucólisis, como cualquier ruta metabólica, solo debe llevarse a cabo cuando las necesidades de la célula lo requieran.
- Niveles altos de ATP: inhiben la glucólisis.
- Niveles altos de AMP y ADP: activan la glucólisis.
Características de la glucólisis:
- Ocurre en el hialoplasma.
- Es una ruta anaeróbica.
- Produce ATP mediante fosforilación a nivel de sustrato.
- Genera poder reductor en forma de NADH.
- Suministra a la célula varios metabolitos intermedios.
Destinos del piruvato originado en la glucólisis
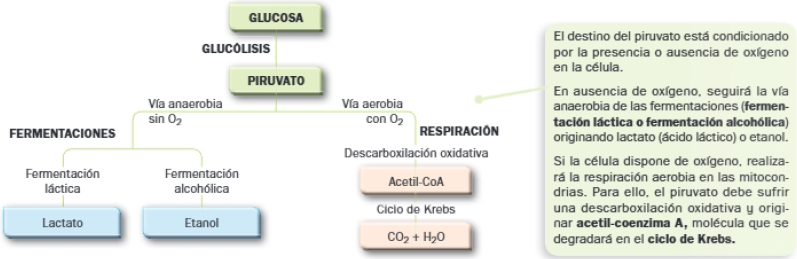
B. Fermentaciones
Los microorganismos aerobios obtienen ATP mediante fosforilación a nivel de sustrato durante las reacciones de la glucólisis; lo mismo ocurre en las células del músculo esquelético cuando no disponen de oxígeno.
La fermentación es un proceso catabólico anaerobio que ocurre en el hialoplasma y en el que se produce una degradación de la materia orgánica a biomoléculas orgánicas más simples, con el fin de obtener energía.
Las fermentaciones constan de dos etapas:
- Etapa 1: Glucólisis, que degrada glucosa a piruvato y, además, se obtienen 2 ATP y 2 NADH.
- Etapa 2: Reducción del piruvato a lactato (fermentación láctica) o a etanol (fermentación alcohólica).
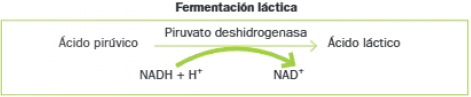
Realizan la fermentación láctica las bacterias lácticas y las células del músculo esquelético en ausencia de oxígeno.
El ácido láctico disminuye el pH y provoca la desnaturalización de las proteínas de la leche, que se vuelven y precipitan. Se obtienen así derivados lácteos como el queso o el yogur.
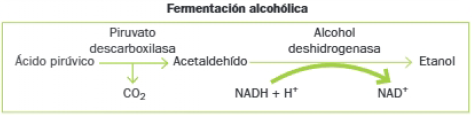
Realizan fermentación alcohólica levaduras del género Saccharomyces, empleadas industrialmente en la obtención de bebidas alcohólicas como el vino o la cerveza, dependiendo de la especie y del proceso. También en la elaboración del pan: actúan sobre los azúcares de las harinas; el CO2 aumenta de volumen de la masa y el etanol se evapora durante la cocción.
Se habla de fermentación heteroláctica cuando se produce una mezcla de lactato y etanol.
Características de las fermentaciones:
- Son rutas catabólicas anaerobias.
- Transcurren por el hialoplasma.
- Son degradaciones incompletas: el producto final es un compuesto orgánico.
- Consumen el NADH de la glucólisis y restauran el NAD+.
- No se obtiene más ATP que el de la glucólisis (2 ATP).
- Son el mecanismo más antiguo de obtención de energía en las células (la atmósfera primitiva carecía de oxígeno).
- Las células anaerobias estrictas (ciertas bacterias) solo pueden obtener energía de los compuestos orgánicos a través de las fermentaciones.
C. Respiración aerobia
Si la célula tiene oxígeno, la degradación del piruvato obtenido en la glucólisis se completa en la mitocondria.
La respiración aerobia puede definirse como el conjunto de procesos catabólicos que ocurren en las mitocondrias, en los cuales se produce la degradación total de las moléculas orgánicas a dióxido de carbono (CO2) y agua (H2O), con obtención de energía en forma de ATP y consumo de oxígeno (O2).
En la respiración aerobia diferenciamos las siguientes etapas:
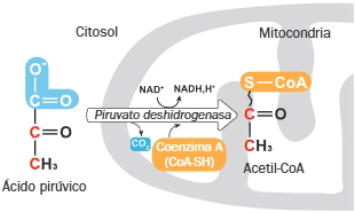
1 Obtención del acetil-CoA
Partiendo del piruvato de la glucólisis, este sufre una descarboxilación oxidativa que origina acetil-CoA, CO2 y NADH. La reacción está catalizada por la enzima piruvato deshidrogenasa.
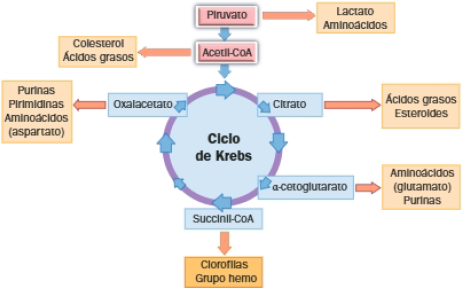
2 Ciclo de Krebs
Es la última etapa de la degradación de glúcidos, lípidos e incluso de muchos aminoácidos.
El ciclo de Krebs es una ruta antibólica que, en la respiración aerobia, realiza la degradación del acetil-CoA a CO2, a la vez que se genera GTP y coenzimas reducidas (NADH y FADH2).
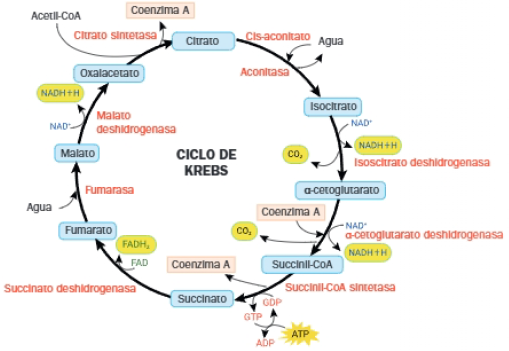
Balance del ciclo de Krebs:
Por cada molécula de acetil-CoA que entra en el ciclo, se obtienen:
- 2 CO2.
- 1 FADH2.
- 3 NADH.
- 1 GTP (=1ATP).
- Se regenera el CoA-SH.
Se ha completado la degradación de la glucosa:
Los 6 carbonos de la glucosa se han desprendido en forma de 6 CO2: 2 en la descarboxilación oxidativa del piruvato y 4 en el ciclo de Krebs.
El CO2 es un residuo de la respiración que es expulsado de las células.
Cadena de transporte electrónico mitocondrial y fosforilación oxidativa
La cadena de transporte también se conoce como cadena respiratoria. Es la última fase de la respiración mitocondrial. En esta fase interviene el oxígeno (O2).
Las coenzimas reducidas NADH y FADH2 generadas en todos los procesos anteriores (glucólisis, descarboxilación oxidativa del piruvato y ciclo de Krebs) ceden los electrones, que han captado en las reacciones de oxidación, a una cadena de transportadores de electrones situada en la membrana mitocondrial interna.
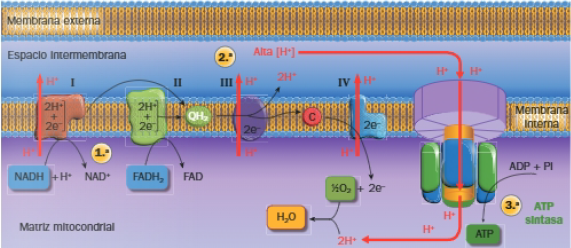
En este proceso podemos diferenciar 3 etapas:
- Etapa 1: NADH y FADH2 ceden electrones a la cadena transportadora de electrones situada en las crestas de la membrana mitocondrial interna. Los componentes de la cadena son proteínas de membrana con grupos prostéticos capaces de aceptar y ceder electrones y están agrupadas en 4 complejos supramoleculares o sistemas; el NADH cede electrones al complejo I y el FADH2 al complejo II.
A través de la cadena de transporte electrónico los electrones llegan hasta el oxígeno, que es el aceptor final de los H+ y los electrones. - Etapa 2: Generación de un gradiente electroquímico. Según la teoria quimiosmótica la energía que los electrones van perdiendo al pasar a través de la cadena, se utiliza para bombear H+ desde la matriz al espacio intermembrana, a través de los complejos I, III y IV, generando un gradiente químico y eléctrico.
- Etapa 3: Síntesis de ATP mediante fosforilación oxidativa. En la membrana mitocondrial interna están ubicadas las ATP sintasas o ATPasas, con un canal en su interior a través del cual pueden retornar los H+ a la matriz mitocondrial. El paso de H+ activa las ATPasas.
Por cada NADH se forman 2,5 ATP; por cada FADH2 se forman 1,5 ATP.
El transporte electrónico y la fosforilación oxidativa están acoplados, pero parte de la energía se pierde en forma de calor. La célula puede utilizar el gradiente electroquímico para la síntesis de ATP, producción de calor, transporte activo y movimiento de cilios y flagelos.

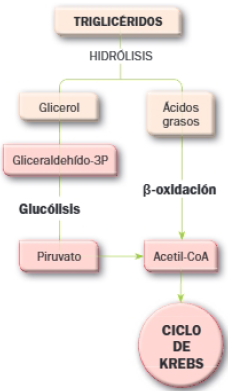
Catabolismo de los lípidos
Los triglicéridos son moléculas de reserva energética a las que la célula puede recurrir para obtener energía. El primer paso es la hidrólisis, de la cual resulta glicerol y tres ácidos grasos. El glicerol se transforma en gliceraldechído-3-fosfato y, de este modo, se incorpora a la glucólisis.
Los ácidos grasos se degradan en la matriz mitocondrial. Su degradación implica tres etapas: activación, entrada en la mitocondria y beta–oxidación.
1 Activación de los ácidos grasos: formación de acil-CoA graso
Consiste en la unión del ácido graso al CoA, formando un acil-CoA graso o ácido graso activado. Tiene lugar en la cara citosólica de la membrana mitocondrial externa o de la membrana del retículo endoplasmático liso.
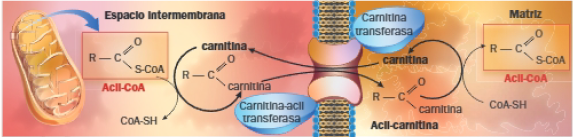
2 Entrada del ácido graso activado en la matriz mitocondrial
Los ácidos grasos de cadena corta (hasta 12 carbonos) atraviesan la bicapa lipídica de las membranas mitocondriales; los ácidos grasos de cadena larga (más de 12 carbonos) necesitan una lanzadera que los introduzca en la matriz mitocondrial: la lanzadera de carnitina, que consume ATP.
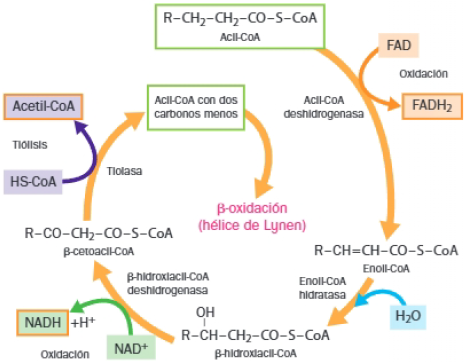
3 Beta-oxidación de los ácidos grasos o hélice de Lynen
Órganos como el hígado o el corazón obtienen el 80% de la energía a través de la beta-oxidación.
La beta-oxidación de los ácidos grasos es un proceso catabólico cíclico mediante el cual los ácidos grasos liberan dos átomos de carbono en forma de acetil-CoA por cada vuelta del ciclo.
2 ANABOLISMO
El anabolismo es el conjunto de rutas metabólicas en las que se sintetizan moléculas complejas a partir de otras más simples, utilizando energía en forma de ATP. Dentro del anabolismo, podemos diferenciar dos tipos:
- Anabolismo autótrofo, que utiliza como fuente de carbono el CO2. Incluye la fotosíntesis y la quimiosíntesis. Ambas se diferencian en la fuente de energía: en la fotosíntesis procede de la luz solar y en la quimiosíntesis de reacciones químicas exotérmicas.
- Anabolismo heterótrofo, que utiliza como fuente de carbono biomoléculas orgánicas.
Fotosíntesis
La vida en el planeta depende de los organismos fotosintéticos, pues estos son capaces de sintetizar materia orgánica de nuevo, a partir de la inorgánica. Por eso constituyen la base en la que se asientan cadenas tróficas de los ecosistemas y constituyen el primer nivel trófico: el de los productores.
La fotosíntesis es el proceso mediante el cual determinadas células captan la energía de la luz solar y la utilizan para sintetizar materia orgánica a partir de la inorgánica.
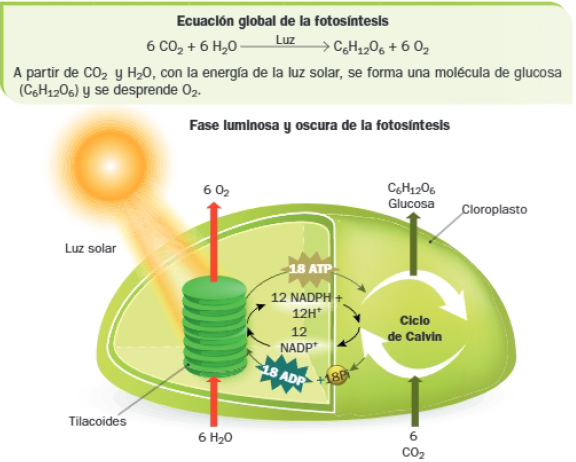
Las plantas verdes, las algas y las cianobacterias realizan la denominada fotosíntesis oxigénica, en la cual se desprende oxígeno; del oxígeno dependen los seres vivos aerobios.
Las bacterias fotosintéticas, como las sulfurosas purpúreas o verdes, realizan la fotosíntesis anoxígena, en la que no se libera oxígeno por que le agente reductor no es el agua, sino compuestos como SH2, en cuyo caso, dependen del azufre (S).
A. Fases de la fotosíntesis
La fotosíntesis oxigénica, llevada a cabo por las plantas, ocurre en los cloroplastos y tiene lugar en dos fases conocidas como fase lumínica y fase oscura.
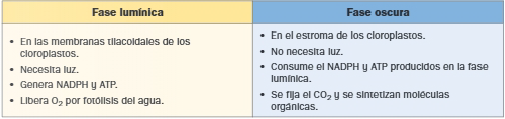
Fase lumínica
La fase lumínica consiste en la absorción de energía luminosa por parte de los pigmentos y su transformación en energía química en forma de ATP y NADPH, acompañada de la liberación de oxígeno.
Las sustancias captadoras de energía de luz son los pigmentos; concretamente, en las plantas son los carotenoides y las clorofilas.
En los tilacoides, los pigmentos están agrupados en dos fotosistemas. Un fotosistema es un complejo multiproteico cuya misión es captar la energía lumínica.
En la fase lumínica se produce un flujo de electrones según en denominado esquema en Z:
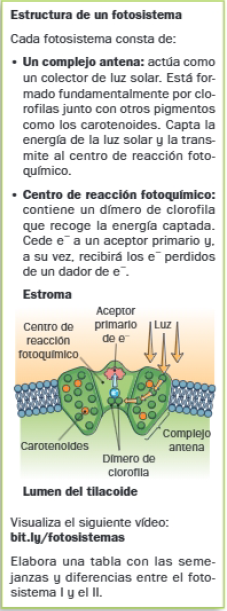
- El PSI absorbe luz, los electrones excitados pasan a través de una cadena de transportadores de electrones hasta el NADP+, conlo que al PSI le faltan electrones, le queda un hueco.
- El PSII absorbe la luz y los electrones excitados pasan por una cadena central de transportadores de electrones hasta el PSI.
- Fotólisis del agua: el PSII necesita electrones, que le son cedidos por el agua.
- Como consecuencia de este proceso, el NADP+ capta H+ y electrones y se transforma en NADPH.
- En el paso del PSII a PSI hay un desprendimiento de energía que es utilizada para bombear H+ desde el estroma al interior del tilacoide, generando un gradiente electroquímico entre ambos espacios, al aumentar la concentración de H+ en el interior el tiolacoide. Al igual que pasaba en la membrana mitocondrial interna, los H+ no pueden atravesar la membrana tilacoidal. Para retornar al estroma, deben hacerlo a través de la ATPsintasa o ATPasa, que se activa y sintetiza ATP.
Transporte electrónico fotosintético no cíclico y cíclico
El flujo de electrones descrito como esquema en Z, desde el agua hasta el NADP+, se conoce transporte de electrones fotosintético no cíclico.
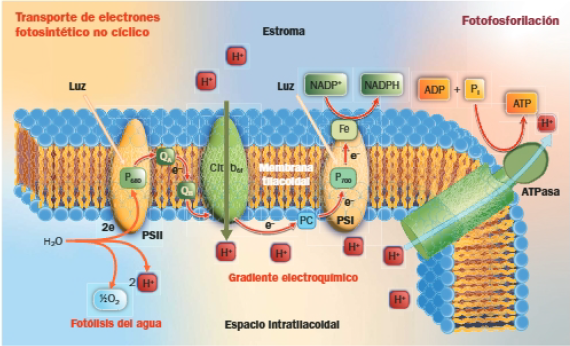
El balance del transporte fotosintético no cíclico es el siguiente:

Los H del agua acaban en el NADPH +H+; el O2 se desprende y se sintetiza ATP.
En ocasiones, la célula necesita ATP, pero no NADPH, entonces recurre al transporte de electrones fotosintético cíclico:
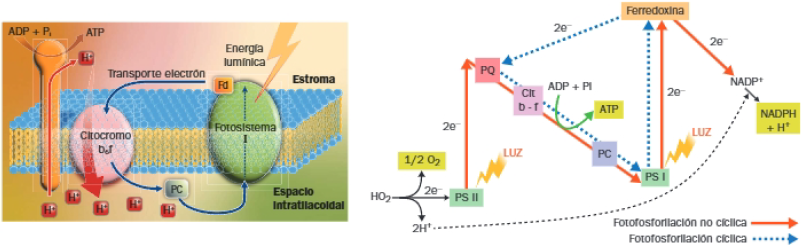
- En el transporte cíclico, solo interviene el PSI y los electrones retornan a él a través de un flujo cíclico: de la ferredoxina, retornan a la PQ y, a través de la cadena central, vuelven al PSI.
- En el paso de los electrones a través de la cadena central de transportadores de electrones se libera energía, utilizada para bombear H+ al interior del tilacoide; el gradiente electroquímico generado activa la ATPasa, al retornar los H+ al estroma a través de ella.
- No se produce NADPH ni O2, tan solo ATP.
Fase oscura de la fotosíntesis: ciclo de Calvin.
La fase oscura consiste en la utilización del NADPH y el ATP, producidos en la fase luminosa para reducir el CO2 a biomoléculas orgánicas.
El ciclo de Calvin se inicia en el estroma del cloroplasto y continúa en el hialoplasma. No necesita luz, pero sí los productos de la fase luminosa: ATP y NADPH.
Las reservas celulares de ATP y NADPH duran solo unos segundos o minutos poque no son moléculas que las células almacenen. Por ello, el ciclo de Calvin depende de que estas moléculas sean aportadas de nuevo por las reacciones luminosas.
Melvin Calvin, junto con Andrew Benson, además de James Bassham, determinó en 1953 la ruta metabólica mediante la cual las plantas convierten el CO2 en azúcares. En 1961, Calvin recibió el premio Nobel por su descubrimiento del denominado ciclo de Calvin, que realizó a través de experimentos en los que exponía algas fotosintéticas a CO2 radiactivo durante periodos de tiempo cada vez más cortos. Utilizando el CO2 radiactivo logró averiguar en qué moléculas se integraba.
El ciclo de Calvin puede dividirse en 3 etapas:
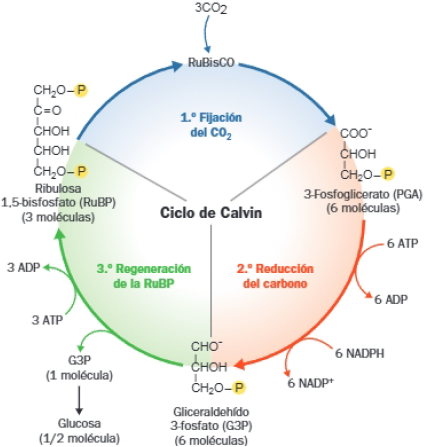
- Fijación del CO2: carboxilación, el CO2 se fija a la ribulosa 1,5-bifosfato (RuBP) mediante la enzima ribulosa-fosfato-carboxilasa o rubisco, la enzima más abundante de la naturaleza.
- Reducción del carbono fijado a gliceraldehído-3-fosfato, lo cual requiere aporte de energía (ATP) y poder reductor (NADPH) generados en la fase lumínica.
- Regeneración de la RuBP, que también requiere aporte energético y la que se destina una parte del gliceraldehído-3-fosfato.
El resto del gliceraldehído-3-fosfato se destina a la síntesis de glúcidos: glucosa, sacarosa y almidón. Finalmente, una parte se usa para la síntesis de lípidos y aminoácidos.
B.Importancia biológica de la fotosíntesis
La importancia de la fotosíntesis puede resumirse en estos puntos fundamentales:
- Transforma la energía de la luz en energía química utilizable por los seres vivos.
- Produce biomoléculas orgánicas, a partir de biomoléculas inorgánicas. Los organismos fotosintéticos son los productores de los ecosistemas, están en la base de las pirámides trófics. La materia orgánica que ellos fabrican sustenta al resto de seres vivos.
- Produce el O2 que hace posible la respiración de los seres aerobios y ha permitido que se forme la capa de ozono (O3) estrastosférico que nos protege de la radiación ultravioleta y hace posible la vida en la superficie de la Tierra.
- Capta CO2 atmosférico, contribuyendo a rebajar sus niveles.
C.Factores que influyen en la fotosíntesis
En la intensidad de la fotosíntesis, influyen factores internos de la propia planta, como el contenido en clorofila o la duración del pigmento en las hojas, y factores externos, como la concentración de CO2, la temperatura, la intensidad de la luz, la abundancia de agua o la concentración de O2.
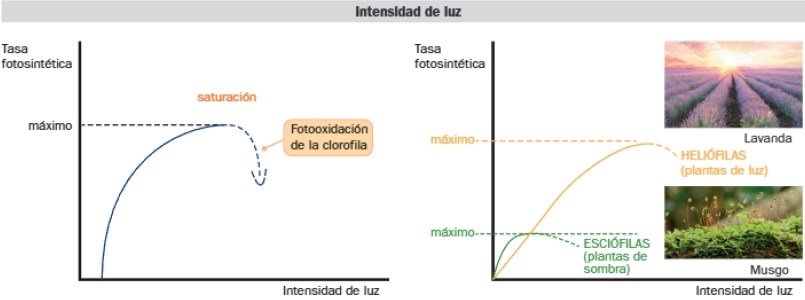
A bajas intensidades lumínicas, un aumento de estas conlleva una mayor tasa fotosintética. Superado el óptimo puede llegar a la saturación e incluso al deterioro de la clorofila por fotooxidación, con la consiguiente disminución de la intensidad de fotosíntesis. El máximo de intensidad de luz varía de unas especies a otras. Las especies esclófilas están adaptadas a bajas intensidades de luz, son plantas de sombra. Las especies hellófilas necesitan intensidades de luz elevadas, son plantas de sol.
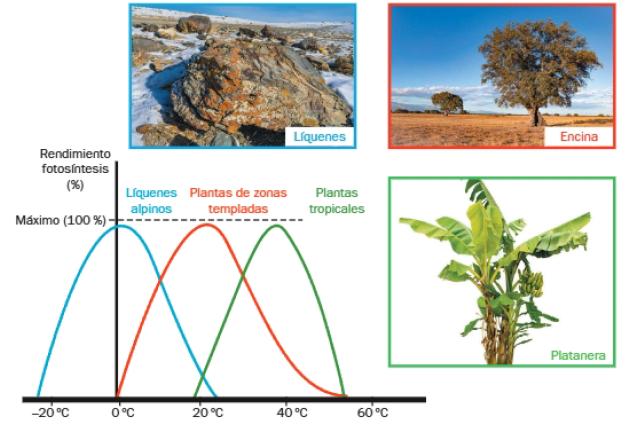
De modo general, un aumento de la temperatura incrementa la velocidad de las reacciones químicas implicadas, hasta un máximo, a partir del cual se produce la desnaturalización de las enzimas implicadas.
La temperatura óptima varía de unas especies a otras. Así, especies de zonas alpinas tienen máximos de rendimiento fotosintético a temperaturas próximas a 0 grados centigrados, mientras que las especies de clima tropical presentan máximos a unos 40 grados centígrados.
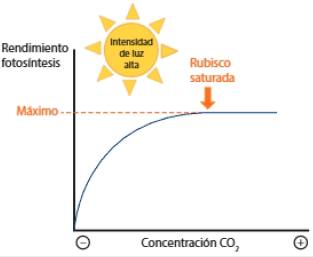
Concentración de CO2
Al representar el rendimiento de la fotosíntesis frente a la concentración de CO2, se obtiene una gráfica que recuerda a la cinética enzimática, al modelo de Michaelis-Menten.
Con intensidades de luz alta, de de modo que se produzca suficiente ATP y NADPH en la fase lumínica, el rendimiento está condicionado a la velocidad con que la rubisco fija el CO2, que actúa como sustrato de la enzima.
La enzima alcanza la velocidad máxima cuando todos los centros activos de todas las moléculas en de enzimas están ocupados. Se dice que la rubisco está saturada.
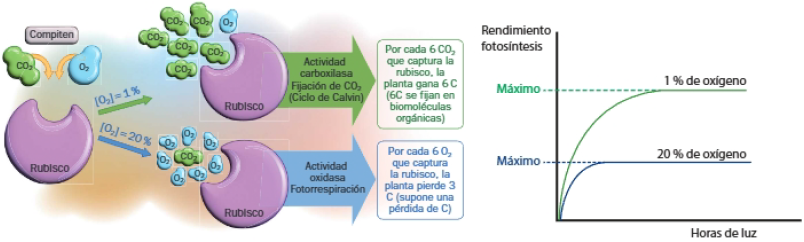
Concentración de O2
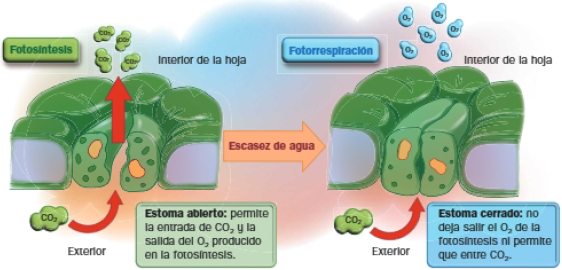
El O2 compite con el CO2 como sustrato de la enzima rubisco. Si es alta, se activa la FOTORESPIRACIÓN: proceso en el que la rubisco, en vez de actuar fijando CO2, captura O2, en un proceso de oxidación que supone una pérdida de átomos de carbono. Así, niveles altos de O2 en la planta suponen un menor rendimiento de la fotosíntesis.
Disponibilidad de agua
La disponibilidad de agua influye en la apertura y cierre de los estromas y, por tanto, en la difusión del CO2 y el O2.
Ante la escasez de agua, los estomas se cierran para evitar pérdidas de la misma. Esto provoca que el CO2 disminuya, mientra que el O2 generado en el interior de la hoja, activando la actividad oxidasa de la rubisco, la fotorrespiración, y disminuyendo el rendimiento fotosintético.
Quimiosíntesis
La quimiosíntesis consiste en la producción de materia orgánica utilizando como fuente de energía la oxidáción de moléculas inorgánicas reducidas, como el ácido sulfhídrico, el hidrógeno gaseoso o el metano, sin necesidad de utilizar la luz solar.
Fases de la quimiosíntesis

Podemos poner algunos ejemplos de bacterias quimiosintéticas, según el sustrato utilizado:
- Bacterias de nitrógeno: Resultan imprescindibles en el ciclo del nitrógeno. Realizan la nitrificación, que consiste en la oxidación del amonio a nitrato. Son bacterias aerobias que utilizan el O2 como agente oxidante. Se trata de microorganismos autótrofos que utilizan el CO2 atmosférico como fuente de carbono. La nitrificación tiene lugar en dos etapas:
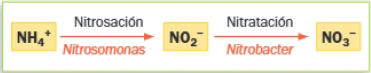
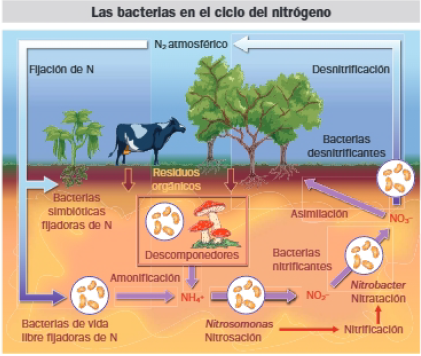
- Bacterias del hierro: Oxidan compuestos ferrosos a férricos. Están presentes en suelos muy ácidos ricos en hierro.
- Bacterias del hidrógeno: Utilizan el hidrógeno molecular y producen metano. Abundan en los pantanos.
- Bacterias incoloras del azufre: Usan H2S como dador de hidrógenos, abundante en aguas residuales como producto de la descomposición de materia orgánica y en aguas hidrotermales. Por ejemplo: arqueas que toleran las altas temperaturas y viven en chimeneas hidrotermales o en géiseres.

Anabolismo heterótrofo
Fotosíntesis y quimosíntesis se engloban dentro del anabolismo autótrofo, puesto que utilizan como fuente de carbono compuestos inorgánicos; pero existen otras rutas metabólicas de síntesis que utilizan como fuente de carbono biomoléculas orgánicas y constituyen el anabolismo heterótrofo. De esas veremos algunos ejemplos representativos, tales como la síntesis de glúcidos como la glucosa (neoglucogénesis) y de glucógeno (glucogenoneogénesis), la síntesis de lípidos (de ácidos grasos o triglicéridos) y la de aminoácidos.
A. Carácter anfibólico del ciclo de Krebs
Hemos visto que el ciclo de Krebs es la etapa final de la degradación de biomoléculas a CO2 y H2O, pero este ciclo participa a la vez en el anabolismo, respondiendo a las necesidades de la célula; por ello se considera una ruta anfibólica.
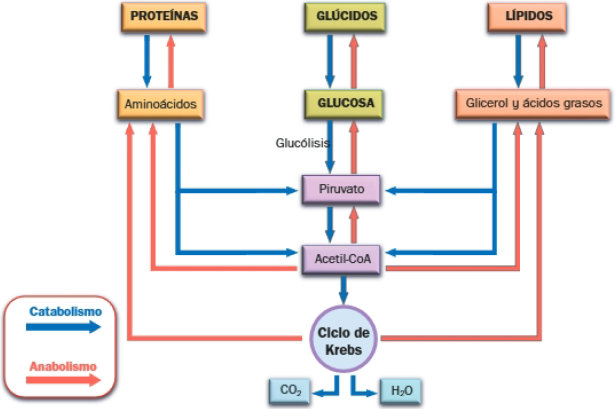
El ciclo de Krebs y el anabolismo
Muchas de las moléculas intermediarias del ciclo de Krebs pueden ser dirigidas hacia procesos de biosíntesis de biomoléculas, es decir, hacia rutas anabólicas.
B.Biosíntesis de glúcidos: gluconeogénesis y glucogenogénesis
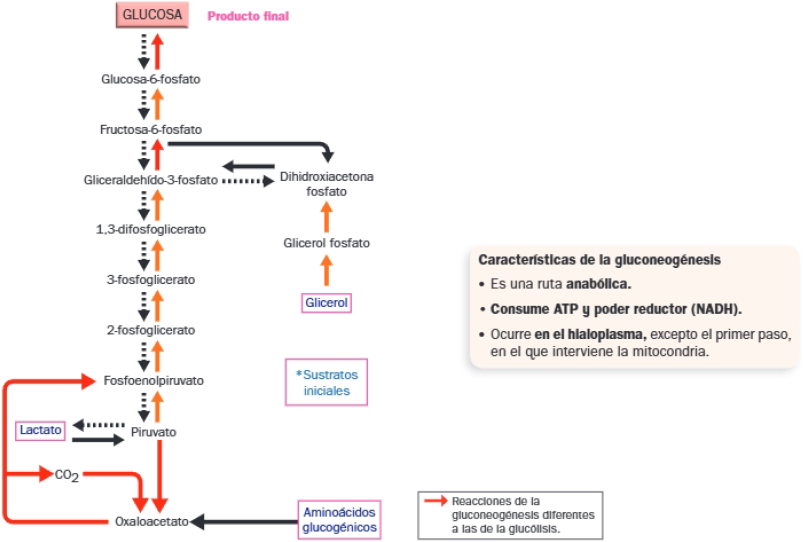
Gluconeogénesis
La gluconeogénesis o neoglucogénesis es la síntesis de glucosa a partir de compuestos orgánicos no glucídicos (aminoácidos, ácido láctico y glicerol).
Ocurre cuando la célula no dispone de la glucosa que necesita. Comparte la mayoría de las reacciones de la glucólisis, pero en sentido contrario, aunque las enzimas que catalizan las reacciones en un sentido (glucólisis) o en el contrario (gluconeogénesis) son diferentes.
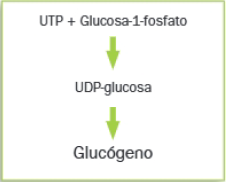
Glucogenogénesis
Es la síntesis de glucógeno a partir de glucosa, con el fin de almacenarla como reserva de energía.
La glucosa puede almacenarse en forma de polisacáridos: almidón en los vegetales y glucógeno en los animales y los hongos.
Para que tenga lugar la síntesis de glucógeno, las glucosas deben ser activadas, en forma de UDP-glucosa; esto se logra al reaccionar la glucosa-1-fosfato con el nucleótido UTP. La síntesis de glucógeno se realiza por adición de unidades de glucosa, siendo la molécula donadora la UDP-glucosa. La reacción está catalizada por la enzima denominada glucógeno sintasa.
La síntesis de glucógeno en los animales tiene lugar en el citoplasma de los hepatocitos (células del hígado) y en las células del músculo esquelético.
C.Lipogénesis: síntesis de triglicéridos
La síntesis de triglicéridos necesita previamente de la síntesis de ácidos grasos.
Síntesis de ácidos grasos
Ocurre en el hialoplasma, donde se localiza el complejo enzimático: ácido graso sintetasa. En las células vegetales fotosintéticas se localiza en el estroma de los cloroplastos.
Consume poder reductor en forma de NADPH.
El sustrato inicial es acetil-CoA: esta es una molécula intermediaria del metabolismo de glúcidos y lípidos. Puede proceder del piruvato de la glucólisis, de la beta oxidación de los ácidos grasos y de aminoácidos, por desanimación o transaminación (eliminación del grupo amino). De este modo, el exceso de cualquiera de la biomoléculas acaba convertido en ácidos grasos y estos almacenados en forma de triglicéridos (grasas) en el tejido adiposo de los animales.
Biosíntesis de triglicéridos
Se produce por esterificación de una molécula de glicerina con tres ácidos grasos, ocurriendo todo esto en el hialoplasma de células hepáticas y adipocitos.
D. Biosíntesis de aminoácidos
Tiene lugar en el citoplasma. Los aminoácidos, ademas de C, H y O, poseen un grupo amino. La síntesis requiere una cadena carbonada a la que se une el grupo amino.
- La cadena carbonada puede originarse a partir de intermediarios de la glucólisis, del ciclo de Krebs o de la denominada ruta de las pentosas fosfato.
- El grupo amino de la mayoría de los aminoácidos procede del glutamato, que transfiere el grupo amino a una cadena carbonada. La glutamina, aminoácido que tiene un grupo amino extra, también puede cederlo a una amplia gama de compuestos.
El proceso de síntesis tiene lugar en dos fases:
- Fase 1: Formación del glutamato a partir del alfa-cetoglutarato del ciclo de Krebs:
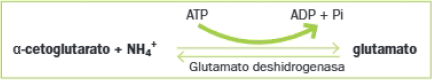
En el caso de la glutamina, se forma a partir del glutamato según la reacción siguiente:
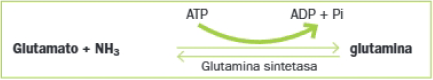
- Fase 2: Transferencia del grupo amino a la cadena carbonada: el glutamato transfiere, mediante reacciones de transaminación, el grupo amino a diferentes esqueletos de carbono para sintetizar aminoácidos. En otros casos, la glutamina cede el grupo amino de la cadena lateral.
Fuente: Biología 2 (Mc Graw Hill).